Algunos ejemplos:
T.1
J17A2
. (2 puntos) a) Defina el concepto de oligoelemento y ponga dos ejemplos nombrando el papel que juegan en el funcionamiento del organismo. (0,75 puntos) b) Defina biomolécula y cite dos ejemplos. (0,5 puntos) c) Nombre tres funciones de las sales minerales. (0,75 puntos)
a)Es un bioelemento que se encuentra en cantidades muy bajas.(0.1%-oligo=bajo).
P.e Na+ y K+ imprescindibles para la transmisión del impulso nervioso, éste último y el Ca+ intervienen en la contracción muscular.
b) Moléculas presentes en la materia viva .P.e Glúcidos son b. orgánicas y sales minerales son b.inorgánicas ,es decir tb las encontramos fuera de los s.v.
c) -Constitución de estructuras duras de protección o sosten P.e en huesos,conchas...
-Funciones fisiológicas,p.e Cationes Na+ y K+ imprescindibles para la transmisión del impulso nervioso, éste último y el Ca+ intervienen en la contracción muscular.
-Mantenimiento de concentraciones osmóticas adecuadas.
J15A2
-En la gráfica adjunta se representa la variación del volumen de una célula en función del tiempo. La célula fue colocada inicialmente en un medio con alta concentración de sales y a los 10 minutos fue transferida a un medio con agua destilada. (2 puntos) a) Proponga una explicación razonada a los cambios de volumen que sufre la célula a lo largo del tiempo. (1 punto) b) Nombre cuatro funciones de las sales minerales. (1 punto)

Ósmosis: paso de agua de un medio más diluido a un medio más concentrado a través de
membranas semipermeables.
Al principio al estar en un medio con mucha concentración, el volumen de la célula disminuye
porque sale agua de su interior. Si se pone en un medio con agua destilada, entra agua al interior
de la célula al ser el medio más diluido y la célula aumenta de volumen.
En el primer caso la célula está en un medio hipertónico y por eso sale agua de su interior.
En el segundo caso está en un medio hipotónico y entra agua al interior de la célula.
b)
− Función esquelética: sustancias minerales precipitadas como fosfato de calcio, carbonato de
calcio que depositado sobre el colágeno constituye los huesos. La sílice forma parte de las
algas diatomeas.
− Sales disueltas como sodio, potasio, calcio, magnesio, cloro, hierro. Estos iones ayudan a
mantener el grado de salinidad.
− Los iones intervienen en el poder tampón o amortiguador del organismo.
− Iones como el hierro forman parte de la hemoglobina.
− El sodio y el potasio intervienen en la transmisión del impulso nervioso.
− El potasio aumenta la turgencia de la célula.
− El yodo forma parte de la tiroxina.
− Las sales minerales pueden estar asociadas a moléculas orgánicas como fosfolípidos,
fosfoproteínas, etc
S13B4
- Razone por qué no es conveniente introducir un pez marino en agua dulce. ¿Cómo se denominan los fenómenos que sufrirían sus células? (1 punto)
La ósmosis es un caso particular de difusión en el que dos disoluciones de diferente concentración y separadas por una membrana semipermeable igualan sus concentraciones(d.isotónicas) al pasar agua de la más diluida (d. hipotónica) a la más concentrada(d.hipertónica).
Así pues al introducir una célula,tejido...en un medio no isotónico respecto de aquellos habrá un movimiento de agua, en este caso sería desde el medio externo(agua dulce-menos concentrado-d.hipotónica) al m. interno produciéndose el fenómeno de turgencia.
S14B2
-Un tejido animal o vegetal se introduce en soluciones de diferentes concentraciones osmóticas: (1 punto) a) ¿Qué ocurriría si la solución utilizada fuera hipotónica? Razone la respuesta. (0,35 puntos) b) ¿Y si la solución utilizada fuera hipertónica? Razone la respuesta. (0,35 puntos) c) Explique con qué propiedad de la membrana plasmática están relacionadas las respuestas de los apartados anteriores. (0,3 puntos)
La ósmosis es un caso particular de difusión en el que dos disoluciones de diferente concentración y separadas por una membrana semipermeable igualan sus concentraciones(d.isotónicas) al pasar agua de la más diluida (d. hipotónica) a la más concentrada(d.hipertónica).
a) Al introducir una célula,tejido...en un medio no isotónico respecto de aquellos habrá un movimiento de agua, en este caso sería desde el medio externo(menos concentrado-d.hipotónica) al m. interno(la célula está mas concentrada-m.hipertónica) produciéndose el fenómeno de turgencia.
b) Al introducir una célula,tejido...en un medio no isotónico respecto de aquellos habrá un movimiento de agua, en este caso sería desde el medio interno(mas concentrado-d.hipertónica) al m. externo (en este caso menos concentrado d. hipotónica)produciéndose el fenómeno de plasmólisis.
c) Como ya se ha indicado en la introducción a la pregunta la ósmosis se realiza a través de una membrana caracterizada por una semipermeabilidad que impide el paso de soluto(sales)

T.2
S19
OPCIÓN A1. Algunas de nuestras prendas de vestir tienen un origen vegetal y están hechas de algodón, que contiene celulosa. Otras, como la seda, son de origen animal. La seda está formada por una proteína que forma láminas β con cadenas que repiten una secuencia de glicina-serina-glicina-alanina-glicina-alanina. (2 puntos)a) Indique, concretando lo máximo posible, qué tipo de biomolécula es la celulosa ¿Qué función tiene en el vegetal de origen? ¿De qué estructura forma parte? (0,5 puntos) b)¿Cómo se llama el enlace que se produce entre los monómeros constituyentes de la celulosa? ¿De qué tipo es? Razónelo. (0,5 puntos)c) ¿Qué es una lámina β? (0,5 puntos)d) ¿Qué tipo de enlace unirá la glicina con serina? Exprese la formación del enlace de una forma general. Cite dos características de dicho enlace. (0,5 puntos
a ) 0,5 puntos:Polisacárido (0,1 puntos). Función estructural (0,2 puntos). Forma parte de la pared celular (0,2 puntos)
b) 0,5 puntos: Es un enlace O-glucosídico (0,2 puntos). Es un enlace tipo β(1->4) dado que une la secuencia de β-D-glucosas. El grupo OH del C anomérico está en posición cis, es decir, hacia arriba(0,3 puntos).
S18B4a,b
. Explique brevemente:
a) ¿En qué se diferencian las aldosas de las cetosas? (0,5 puntos) b) ¿Qué es un carbono asimétrico y qué repercusión tiene su presencia en estas moléculas? (0,5 puntos)
a)Aldosa: Glúcido monosacárido cuyo grupo carbonilo es un grupo aldehido –CHO (0,25 puntos) Cetosa: Glúcido monosacárido cuyo grupo carbonilo es un grupo cetona –CO- (0,25 puntos)
 b)Carbono que se enlaza a cuatro grupos atómicos diferentes,las moléculas que lo poseen tendrán estereoisomería
b)Carbono que se enlaza a cuatro grupos atómicos diferentes,las moléculas que lo poseen tendrán estereoisomería

-Tema de desarrollo corto: Glúcidos. (3 puntos) a) Definición. (0,5 puntos) b) Características químicas. (0,5 puntos) c) Funciones. (1 punto) d) Clasificación. (1 punto)
-Un tejido animal o vegetal se introduce en soluciones de diferentes concentraciones osmóticas: (1 punto) a) ¿Qué ocurriría si la solución utilizada fuera hipotónica? Razone la respuesta. (0,35 puntos) b) ¿Y si la solución utilizada fuera hipertónica? Razone la respuesta. (0,35 puntos) c) Explique con qué propiedad de la membrana plasmática están relacionadas las respuestas de los apartados anteriores. (0,3 puntos)
La ósmosis es un caso particular de difusión en el que dos disoluciones de diferente concentración y separadas por una membrana semipermeable igualan sus concentraciones(d.isotónicas) al pasar agua de la más diluida (d. hipotónica) a la más concentrada(d.hipertónica).
a) Al introducir una célula,tejido...en un medio no isotónico respecto de aquellos habrá un movimiento de agua, en este caso sería desde el medio externo(menos concentrado-d.hipotónica) al m. interno(la célula está mas concentrada-m.hipertónica) produciéndose el fenómeno de turgencia.
b) Al introducir una célula,tejido...en un medio no isotónico respecto de aquellos habrá un movimiento de agua, en este caso sería desde el medio interno(mas concentrado-d.hipertónica) al m. externo (en este caso menos concentrado d. hipotónica)produciéndose el fenómeno de plasmólisis.
c) Como ya se ha indicado en la introducción a la pregunta la ósmosis se realiza a través de una membrana caracterizada por una semipermeabilidad que impide el paso de soluto(sales)

S19
OPCIÓN A1. Algunas de nuestras prendas de vestir tienen un origen vegetal y están hechas de algodón, que contiene celulosa. Otras, como la seda, son de origen animal. La seda está formada por una proteína que forma láminas β con cadenas que repiten una secuencia de glicina-serina-glicina-alanina-glicina-alanina. (2 puntos)a) Indique, concretando lo máximo posible, qué tipo de biomolécula es la celulosa ¿Qué función tiene en el vegetal de origen? ¿De qué estructura forma parte? (0,5 puntos) b)¿Cómo se llama el enlace que se produce entre los monómeros constituyentes de la celulosa? ¿De qué tipo es? Razónelo. (0,5 puntos)c) ¿Qué es una lámina β? (0,5 puntos)d) ¿Qué tipo de enlace unirá la glicina con serina? Exprese la formación del enlace de una forma general. Cite dos características de dicho enlace. (0,5 puntos
a ) 0,5 puntos:Polisacárido (0,1 puntos). Función estructural (0,2 puntos). Forma parte de la pared celular (0,2 puntos)
b) 0,5 puntos: Es un enlace O-glucosídico (0,2 puntos). Es un enlace tipo β(1->4) dado que une la secuencia de β-D-glucosas. El grupo OH del C anomérico está en posición cis, es decir, hacia arriba(0,3 puntos).
 |
| Disacarido que forma la celulosa. |
. Explique brevemente:
a) ¿En qué se diferencian las aldosas de las cetosas? (0,5 puntos) b) ¿Qué es un carbono asimétrico y qué repercusión tiene su presencia en estas moléculas? (0,5 puntos)
a)Aldosa: Glúcido monosacárido cuyo grupo carbonilo es un grupo aldehido –CHO (0,25 puntos) Cetosa: Glúcido monosacárido cuyo grupo carbonilo es un grupo cetona –CO- (0,25 puntos)
 |
| El 2º C es el asimétrico o quiral,se suele representar con un superíndice asterisco,los grupos atómicos están en diferente color. |

 |
| Todos pertenecen al mismo grupo carbonílico,en este caso aldehído-aldosas-.Si cambian la disposición de todos los grupos -OH-grupo alcohol o hidroxilo-son enantiómeros(imagenes especulares),si cambia uno sólo serían epímeros y en cualquier caso todos son estereoisómeros( el término dias... se reserva a aquellos estereo... que no son enatio... |
-Tema de desarrollo corto: Glúcidos. (3 puntos) a) Definición. (0,5 puntos) b) Características químicas. (0,5 puntos) c) Funciones. (1 punto) d) Clasificación. (1 punto)
a) Son polihidroxialdehídos o polihidroxicetonas de fórmula empírica general (CH2O)n
b) Dependiendo del tipo de g. tenemos los monos. que no son hidrolizable , sí reductores .Los disacáridos y polis. son hidrolizables .Los d. son reductores excpto la sacarosa y los poli son poco red(tienen un gr. -OH del carbono con la f. carbonilo libre)
c)

b) Dependiendo del tipo de g. tenemos los monos. que no son hidrolizable , sí reductores .Los disacáridos y polis. son hidrolizables .Los d. son reductores excpto la sacarosa y los poli son poco red(tienen un gr. -OH del carbono con la f. carbonilo libre)
c)
d) Los g. se pueden clasificar en osas u ósidos según estén constituidos por una o varias unidades glucídicas (osas).Las osas o monosacáridos, entre los que destacan la glucosa o la fructosa, pueden también tener derivados como el manitol.
Los ósidos serán disacáridos si están constituidos por dos osas tris.,por tres ,oligo por algunas y poli. por muchas(más de 10)
Entre los di. tenemos la sacarosa o la lactosa y entre los poli según sean homopoli o heteropoli tendremos almidón,glucógeno o celulosa en el primer caso, quitina p.e en el 2º caso.
S18B4 a,b o S12A5a,b
Cuestión .- Explique brevemente:
a) ¿En qué se diferencian las aldosas y las cetosas? (0.5 puntos).
b) ¿Qué repercusión tiene la presencia de un carbono asimétrico? (0.5 puntos).
a) Las aldosas son osas (monosac.) con f. aldehído(C.1) , las cetosas con f. cetona (en C. 2).
b) Un carbono asimétrico es aquél que está unido a cuatro radicales o

Los ósidos serán disacáridos si están constituidos por dos osas tris.,por tres ,oligo por algunas y poli. por muchas(más de 10)
Entre los di. tenemos la sacarosa o la lactosa y entre los poli según sean homopoli o heteropoli tendremos almidón,glucógeno o celulosa en el primer caso, quitina p.e en el 2º caso.
S18B4 a,b o S12A5a,b
Cuestión .- Explique brevemente:
a) ¿En qué se diferencian las aldosas y las cetosas? (0.5 puntos).
b) ¿Qué repercusión tiene la presencia de un carbono asimétrico? (0.5 puntos).
a) Las aldosas son osas (monosac.) con f. aldehído(C.1) , las cetosas con f. cetona (en C. 2).
b) Un carbono asimétrico es aquél que está unido a cuatro radicales o
sustituyentes distintos. Su existencia permite diferenciar los llamados estereoisómeros (moléculas con la misma fórmula molecular pero diferente disposición atómica espacial):
Asimismo, los monosacáridos que poseen uno o más carbonos asimétricos presentan isomería
óptica. Este tipo de isomería se manifieste cuando las moléculas están en disolución y pueden
hacer girar cierto ángulo el plano en que vibra una haz de luz polarizada cuando éste atraviesa
la solución.
Cite el papel que le parezca más relevante de los desempeñados por los siguientes glúcidos: glucosa,ribosa, celulosa, almidón, glucógeno (1 punto).
c) Ribosa y celulosa tendrían f. estructural . El resto energética.
J17B5
Cite el papel que le parezca más relevante de los desempeñados por los siguientes glúcidos: glucosa,ribosa, celulosa, almidón, glucógeno (1 punto).
c) Ribosa y celulosa tendrían f. estructural . El resto energética.
J17B5
En relación con los glúcidos:(2 puntos)
a)(0,9 puntos)
a.1.
Indique cuál de los siguientes compuestos son monosacáridos, disacáridos o polisacáridos:sacarosa, fructosa, almidón, lactosa, celulosa y glucógeno.(0,6 puntos)
a.2.Indique en qué tipo de organismos se encuentran los polisacáridos indicados en el apartado anterior.(0,3 puntos)
b)Diga cuál es la función principal de los polisacáridos indicados en el apartado a) de esta cuestión.(0,3 puntos)
c)Cite dos monosacáridos que conozca y que no se encuentren en la relación
incluida en el apartado a) e indique dónde se pueden encontrar y qué función desempeñan.(0,8 puntos
a) monos. :F;Dis: S,L
a2) Ani:G ;Veg y bacterias:A(tb protistas),C;
b) A y G de reserva energética , la C. f. estructural.
c) Glucosa: libre en p.e uvas(o procedente de hidrólisis de polisac.) tendrá f. energética. Tb unida a otras osas plq tendrá f. estructural,galactosa: unida al glucosa para formar lactosa
Otros:ribosa,desxi....
a2) Ani:G ;Veg y bacterias:A(tb protistas),C;
b) A y G de reserva energética , la C. f. estructural.
c) Glucosa: libre en p.e uvas(o procedente de hidrólisis de polisac.) tendrá f. energética. Tb unida a otras osas plq tendrá f. estructural,galactosa: unida al glucosa para formar lactosa
Otros:ribosa,desxi....

T.3 LÍPIDOS
S19
OPCIÓN B 1. En relación con la figura adjunta, conteste las siguientes cuestiones: (2 puntos) a) ¿Qué tipo de biomolécula representa? (0,2 puntos) b) Indique el nombre de los compuestos incluidos en los recuadros 1 y 2. (0,2 puntos) c) Identifique el tipo de enlace que se establece entre ellos. (0,3 puntos) d) En qué estructura celular se encuentra presente este tipo de biomolécula. (0,2 puntos) e) Si exponemos un gran número de estas moléculas a un medio acuoso ¿Cuál sería su comportamiento? Razone la respuesta. (0,5 puntos) f) Si sustituyéramos el compuesto “4” por otro similar al compuesto “3”, ¿Cambiarían en algo las propiedades de la biomolécula? Razone la respuesta. (0,6 puntos) S18A5c
a) 0,2 puntos: Fosfoglicérido. b) 0,2 puntos: 1 glicerol, 2 ácidos grasos (0,1 puntos cada una). c) 0,3 puntos: enlace éster. d) 0,2 puntos: membrana plasmática. e) 0,5 puntos: formación de micelas o bicapas por su comportamiento anfipático. f) 0,6 puntos: Si. El elemento 3 es un ácido graso saturado y el elemento 4 es un ácido graso insaturado. Los ácidos grasos saturados, debido a la ausencia de los dobles enlaces y su disposición espacial, tienen un punto de fusión mayor, por lo que la nueva molécula tendría un punto de fusión más elevado.
Diferencias entre ácidos grasos saturados e insaturados. (0,5 p)
-Los a.g.s son cadenas hidrocarbonadas con enlaces sencillos y tienen puntos de fusión altos plq son sólidos a Tº ambiente. Ej. ac. palmítico Otros alimentos tocino,aceite de coco...)
-Los a.g.i tienen algún enlace doble y puntos de fusión más bajos siendo líquidos a Tª ambiente. Ej.ac.oléico.Los hay Cis(aceites vegetales) y Trans(margarina)
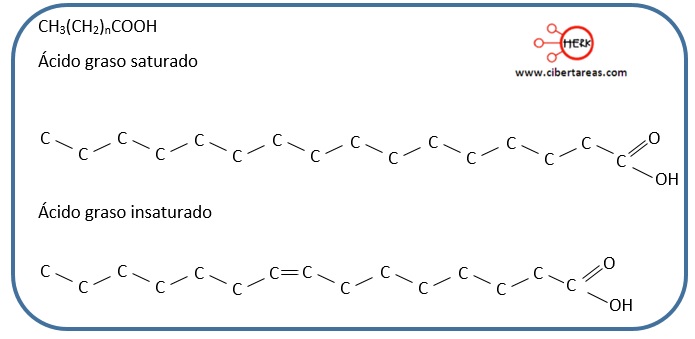
J18A2
En la dieta mediterránea interesa disminuir la grasa saturada y aumentar la grasa poli y monoinsaturada. Una dieta restrictiva en grasa, hace disminuir el colesterol HDL. (2 puntos) a) ¿A qué tipo de biomoléculas pertenecen el colesterol y las grasas? (0,25 puntos) b) ¿Qué significa grasa saturada, monoinsaturada y poliinsaturada. (0,5 puntos) c) ¿En qué consiste el proceso denominado saponificación? (0,75 puntos) d) ¿Qué función desempeña el colesterol en las células animales? (0,5 puntos)
a) Lípidos.
b) Grasas saturadas contienen ácidos grasos que no presentan dobles enlaces entre los carbonos de la cadena hidrocarbonada. Grasas monoinsaturadas son ácidos grasos que contienen un doble enlace.(ej.los ω9 como el aceite de oliva) Polinsaturadas son ácidos grasos con varios dobles enlaces.(Ej los ω 3 y los ω 6 como en los frutos secos y pescado para el primer caso y el aceite de girasol el último)

c) Saponificación es la formación de jabones mediante la reacción química entre un triglicérido y una base (NaOH o KOH). En la reacción partiendo de un triglicérido se forman tres moléculas de las sales sódicas o potásicas correspondientes a los ácidos grasos y se libera glicerina.

d) El colesterol,lípido no saponificable con una función alcohol,entre otras funciones está la estructural concretamente forma parte de las membranas celulares de los animales, regula su fluidez.

S16A1
Tema de desarrollo corto: lípidos. (3 puntos)
a) Explicar qué es un ácido graso. Diferencias entre ácidos grasos saturados e insaturados. Poner un
ejemplo. (1 punto)
b) Diferencias y semejanzas entre triglicéridos y fosfoglicéridos. (1 punto)
c) Indicar cuatro funciones de los lípidos. Poner ejemplos. (1 punto)
a)Es una biomolécula formada por una larga cadena hidrocarbonada lineal, de diferente longitud o número de átomos de carbono, en cuyo extremo hay un grupo carboxilo. Es decir son ácidos orgánicos de cadena larga.

b) S.Químicas:constituidos por a.grasos son saponificables.
D.Químicas: los fosfo. tienen unido un grupo fosfato a un -OH del glicerol y a aquel un aminoalcohol.Sólo dos á. grasos a los otros -OH del glicerol,en el triglicérido son tres.
D.funcionales: Los tri.tienen f. energética,los f.la tienen estructural.

c) Funciones:
-Energética:ciertos lípidos se utilizan como "biocombustibles".P.E los triglicéridos
-Estructural:ciertos lípidos se utilizan para formar otras biomoléculas u orgánulos.P.e fosfolípidos
-Fisiológicas:ciertos lípidos se usan como aislantes térmicos o como protección.Ej grasas en los dos casos , ceras en el último.
-Reguladoras:ciertos lípidos son precursores de hormonas(h.sexuales,suprarrenales) y vitaminas(carotenoides precursores de vit A)
S17A1
1. En relación a los lípidos: (2 puntos)
a) Tipo de reacción que une los ácidos grasos con la glicerina. Indicar el nombre de la reacción
química que resulta. Hacer un esquema. (0,75 puntos)
b) Escribir cuatro funciones de los lípidos indicando un ejemplo de lípido para cada función.
(0,75 puntos)
c) Diferencias entre ácidos grasos saturados e insaturados. (0,5 puntos)
a)Esterificación: ác. grasos + glicerina----> éster(acilglicérido) + agua.

b)Ver S16A1
c)Ver S16A1
J16A5
Forme un triacilglicérido con las siguientes moléculas: (2 puntos)

a) ¿Cómo se llama el enlace que se forma? (0,5 puntos)
El enlace es de tipo éster.Se forma entre los OH de la glicerina y el HOOC- de los ácidos grasos y se desprenden tres moléculas deagua.
b) Además del triacilglicérido, ¿qué otra sustancia obtendremos? (0,5 puntos)
Agua
c) ¿En qué se diferencia un triglicérido de un fosfoglicérido? (1 punto)
Ver S16A1
J14B1
Tema de desarrollo corto: Lípidos. (3 puntos)
a) Características de los lípidos. (0,5 puntos)
b) Ácidos grasos saturados e insaturados. (0,5 puntos)
c) Propiedades físicas y químicas de los ácidos grasos. (1 punto)
d) Diferencia entre acilglicéridos y ceras. (1 punto)
a)Los lípidos constituyen un grupo muy heterogéneo de sustancias químicas, tanto desde el punto de vista estructural como de las funciones que realizan. Están compuestos, básicamente, por carbono e hidrógenoy la mayoría, también presentan oxígeno, pero en proporciones muy bajas. Algunos lípidos, además contienen fósforo, nitrógeno y azufre.
Cumplen dos características distintivas:
− Son insolubles en agua y en otros disolventes polares.
− Son solubles en disolventes orgánicos, es decir disolventes no polares como el éter.
Otras como ser untuosos al tacto,aspecto graso con cierto brillo...
Los lípidos desempeñan funciones biológicas muy variadas. Algunos lípidos almacenan y transportan la energía necesaria para las reacciones metabólicas, otros son componentes estructurales fundamentales en las membranas celulares, relacionados, por ejemplo, con el reconocimiento de las células, o también forman cubiertas externas en los vegetales. Además, hay otros lípidos que, aun en pequeñas cantidades tienen una gran actividad biológica, como es el caso de algunas hormonas y de algunas vitaminas.
T4 Proteínas
S12
Explique brevemente: d) Estructura primaria de las proteínas. (0,5 puntos)
La E.1aria corresponde a la secuencia de aminoácidos de las proteínas. Indica que aminoácidos lo componen y el orden en que se disponen en la cadena. El extremo inicial es aquel que presenta el aminoácido con el grupo amino libre y como extremo final tienen el aminoácido con el grupo carboxilo libre. S19
OPCIÓN A1. Algunas de nuestras prendas de vestir tienen un origen vegetal y están hechas de algodón, que contiene celulosa. Otras, como la seda, son de origen animal. La seda está formada por una proteína que forma láminas β con cadenas que repiten una secuencia de glicina-serina-glicina-alanina-glicina-alanina. (2 puntos)c) ¿Qué es una lámina β? (0,5 puntos)d) ¿Qué tipo de enlace unirá la glicina con serina? Exprese la formación del enlace de una formageneral. Cite dos características de dicho enlace. (0,5 puntos
-Proteinas. Aminoácidos y sus propiedades. Enlace peptídico. Estructuras de las proteínas. Propiedades y clasificación de las Proteinas.
Los aminoácidos son los constituyentes o monómeros que forman las proteínas. Se
trata de compuestos de bajo peso molecular, químicamente formados por carbono,
oxígeno, hidrógeno y nitrógeno. Químicamente se caracterizan por poseer un
grupo carboxilo (-COOH), un grupo amino (-NH2), y una cadena lateral o grupo
R, todos ellos covalentemente unidos a un átomo de carbono denominado α. Los
grupos R son los que determinan las propiedades de los aminoácidos, tales, como la
estructura, tamaño y carga eléctrica.
NH2 -- CH --COOH
R
Los aminoácidos se clasifican en cuatro familias atendiendo a las propiedades de
sus cadenas laterales, en particular a su polaridad. Ésta varía mucho, desde los
grupos totalmente no polares o hidrofóbicos hasta los muy polares e hidrofílicos.
En cuanto a sus propiedades, en general, los aminoácidos son compuestos sólidos,
cristalinos, de elevado punto de fusión, solubles en agua, con actividad óptica y
comportamiento químico anfótero.
El enlace peptídico es un enlace covalente que tiene lugar entre el grupo carboxilo
de un aminoácido y el grupo amino de otro, con la formación de una molécula de
agua. De forma semejante se pueden unir tres aminoácidos mediante dos enlaces
peptídicos formando un tripéptido, cuatro para formar un tetrapéptido, etc. Cuando
se unen varios aminoácidos tenemos un oligopéptido y cuando se unen muchos, un
polipéptido.

La composición y forma de una proteína viene definida por cuatro estructuras,
éstas tienen un carácter jerarquizado, es decir, implican unos niveles o grados de
complejidad creciente que dan lugar a los cuatro tipos de estructuras: primaria,
secundaria, terciaria y cuaternaria.
La estructura primaria de una proteína es la secuencia lineal de los aminoácidos
que contiene, es decir, el número y el orden en el que se encuentran. Los enlaces
peptídicos entre los aminoácidos mantienen estable esta estructura.
La estructura secundaria de una proteína se refiere a la ordenación regular y
periódica en el espacio de la cadena polipeptídica a lo largo de una dirección.
Puede decirse también, que es la disposición de la estructura primara en el espacio
y que es consecuencia directa de la libre capacidad de giro del carbono alfa.
Existen dos modelos o tipos de estructuras secundarias:
- Hélice α .
- Lámina β .
La estructura terciara de una proteína informa de la disposición de la estructura
secundaria en el espacio y, por tanto, del tipo de conformación tridimensional que posee. Las conformaciones más frecuentes que adoptan las proteínas son la
globular y la filamentosa. Las funciones biológicas que realizan las proteínas
dependen de la estructura terciaria que éstas poseen.
La estructura cuaternaria de una proteína informa que ésta está compuesta de
más de una cadena polipeptídica, y hace referencia al modo en que se asocian las
cadenas o subunidades para constituir la proteína activa. (Ej. Hemoglobina). Las
distintas subunidades se unen mediante puentes de hidrógeno y puentes disulfuro.
Las propiedades de las proteínas dependen sobre todo o principalmente de las
características de las cadenas laterales de los aminácidos que las componen y, por
tanto, de su capacidad de reaccionar con otros radicales o con el medio que les
rodea. Las principales propiedades son:
- Solubilidad: La solubilidad de estas moléculas depende:
- del tamaño y forma de la molécula,
- de la disposición de los restos,
- del contenido de electrólitos y del pH del medio.
- Especificidad: Existe de función y de especie.
- Desnaturalización: La mayor parte de las proteínas se pliegan espontáneamente,
adquiriendo la conformación nativa más estable, aunque esta conformación puede
alterarse. La desnaturalización implica la desaparición de las estructuras terciaras
y secundarias originales debido a la ruptura de las interacciones débiles que las
mantienen. En este punto permanece intacta la estructura primaria, la
desnaturalización no afecta a los enlaces peptídicos. La rotura de los puentes de
hidrógeno se puede producir por medios físicos (como radiaciones UVA, calor...) o
químicos (aumentos de pH, disolventes orgánicos..)
Clasificación de las proteínas: atendiendo a su composición se dividen en 2 clases:
simples y conjugadas. Las proteínas simples producen únicamente aminoácidos
cuando se hidrolizan; las conjugadas poseen una parte proteica y una parte no
proteica denominada grupo prostético.
-El DNA (ADN): composición química y estructura (relacione la estructura con la transmisión de la información genética).
El ADN es una biomolécula orgánica compuesta por C, H, O, N y P, que se define
químicamente como un polinucleótido, porque está formado por la repetición de unidades
moleculares llamadas nucleótidos (desoxirribobucleótidos). Su función está relacionada con
el almacenamiento y transmisión de la información hereditaria, constituyendo así la base
molecular de la herencia.
El ADN químicamente está compuesto por tres moléculas diferentes:
1) Una pentosa, que es la desoxirribosa.
2) Una base nitrogenada. Existen dos tipos:
- Púricas: adenina(A) y guanina (G).
- Pirimidínicas: citosina (C), timina (T).
3) Una molécula de ácido fosfórico.
La unión de una pentosa con una base nitrogenada por el carbono 1´ de la pentosa se
denomina nucleósido y la unión de un nucleósido a una molécula de ácido fosfórico a través
del carbono 5´ de la pentosa se denomina nucleótido. La unión de varios nucleótidos por
enlace 5´-3´ fosfodiéster da lugar a un ácido nucleico, que por ello también se denomina
polinucleótido. El encadenamiento de los nucleótidos para formar un ácido nucleico se
realiza siempre mediante el ácido fosfórico, que se une al carbono 3´ de la pentosa del
nucleótido siguiente. Esta molécula tiene dos extremos: un extremo 3´ y un extremo 5´.
El modelo estructural de la doble hélice del ADN, propuesto por Watson y Crick en 1953,
abrió el camino hacia la comprensión de cómo podría desempeñar esta molécula sus
funciones, almacenando y transfiriendo la información genética.
El modelo propone los siguientes aspectos estructurales:
- La existencia de dos cadenas polinucleotídicas dextrógiras arrolladas en forma de hélice
alrededor de un mismo eje constituyendo así una doble hélice. Ambas cadenas o hebras son
antiparalelas, es decir, sus puentes fosfodiéster 3´-5´internucleotídicos van en direcciones
opuestas, una va en sentido 3¢®5¢ y la otra en sentido 5¢®3¢ .
- Las bases púricas y pirimidínicas de cada una de las cadenas o hebras están apiladas en el
interior de la duplohélice, con sus planos paralelos entre sí, y perpendiculares al eje de la
doble hélice. Las bases de una cadena están apareadas mediante puentes de hidrógeno con las
bases de la otra cadena. Los pares permisibles son A-T y G-C.
- Las dos cadenas antiparalelas de la duplohélice no son idénticas en secuencia ni en
composición. En vez de ello, son complementarias entre sí (siempre que en una cadena hay
A, en la otra T, y viceversa).
- Las bases se hallan apiladas a una distancia de 0,34 nm de centro a centro, o lo que es lo
mismo, es la distancia que separa cada par de bases. En cada vuelta completa de la
duplohélice hay exactamente 10 nucleótidos, lo que corresponde a la distancia secundaria
repetida de 3,4 nm. La doble hélice tiene unos 2,0 nm de diámetro.
- Las bases relativamente hidrófobas están situadas en el interior de la hélice y los restos
polares de la pentosa y grupos fosfato cargados negativamente están en la periferia, expuestos al
agua, formando el esqueleto externo de la duplohélice.
- La duplohélice resulta estabilizada no sólo por puentes de hidrógeno entre los pares de bases
complementarias, sino también por interacciones electrónicas entre las bases apiladas, así
como por acciones hidrófobas recíprocas.
En cuanto a la relación estructura-función:
En cuanto a la relación estructura-función:
Cuando Watson y Crick propusieron en 1953 la hipótesis de la estructura molecular en doble
hélice del ADN, surgieron una hipótesis para explicar cómo se encuentra codificada la
información genética. Según ésta, la información genética está contenida en la secuencia de
los nucleótidos (A, T, G, C) de la molécula de ADN( ya que el resto de componentes de los nucleótidos no varían) que determina la secuencia de los aminoácidos en las proteínas.
Por otra parte la replicación es un proceso de autoduplicación del ADN que tiene lugar durante el período
de síntesis del ciclo celular o fase S de la interfase, y se caracteriza porque a partir de una
molécula de ADN, se forman dos iguales a ella e idénticas entre sí. Aunque se han propuesto
varias hipótesis para explicar el mecanismo de este proceso, es, sin embargo, la hipótesis
semiconservativa propuesta por Watson y Crick y demostrada experimentalmente por
Meselson y Stahl en 1957 la de mayor aceptación actualmente. La replicación es
semiconservativa porque las dos cadenas de nucleótidos que forman la doble hélice de ADN
se conservan y sirven de molde para la síntesis de dos hebras complementarias. Por tanto, la
replicación da como resultado dos moléculas de ADN, en las que cada una de ellas se
conserva una cadena antigua, y la otra es nueva.
Son pues las dos características implícitas,la varabilidad de las bases nitrogenadas y la complementariedad entre las cadenas de ADN las que relacionan la estructura del ADN con su función de portador de la información genética.
Son pues las dos características implícitas,la varabilidad de las bases nitrogenadas y la complementariedad entre las cadenas de ADN las que relacionan la estructura del ADN con su función de portador de la información genética.
-ADN. Nucleótidos y nucleósidos. Estructura primaria y secundaria. Los ácidos nucleicos como los compuestos químicos que contienen la información genética.
El ADN es una biomolécula orgánica compuesta por C, H, O, N y P, que se define
químicamente como un polinucleótido, porque está formado por la repetición de unidades moleculares llamadas nucleótidos (desoxirribobucleótidos). Su función está relacionada con el almacenamiento y transmisión de la información hereditaria, constituyendo así la base molecular de la herencia.
El ADN químicamente está compuesto por tres moléculas diferentes:
1) Una pentosa, que es la desoxirribosa.
2) Una base nitrogenada. Existen dos tipos:
- Púricas: adenina(A) y guanina (G).
- Pirimidínicas: citosina (C), timina (T).
3) Una molécula de ácido fosfórico.
La unión de una pentosa con una base nitrogenada por el carbono 1´ de la pentosa se denomina nucleósido y la unión de un nucleósido a una molécula de ácido fosfórico a través del carbono 5´ de la pentosa se denomina nucleótido. La unión de varios nucleótidos por enlace 5´-3´ fosfodiéster da lugar a un ácido nucleico, que por ello también se denomina polinucleótido. El encadenamiento de los nucleótidos para formar un ácido nucleico se realiza siempre mediante el ácido fosfórico, que
se une al carbono 3´ de la pentosa del nucleótido siguiente. Esta molécula tiene dos extremos: un extremo 3´ y un extremo 5´. La estructura primaria del ADN está constituida por la secuencia de los nucleótidos en la cadena. Si se considera el elevado número de nucleótidos
existentes en una hebra de ADN y las innumerables combinaciones posibles, se puede comprender cómo a través de la secuencia de las bases nitrogenadas es posible estructurar una determinada información (el llamado mensaje biológico),de la misma manera que con 26 signos o letras está estructurada la información intelectual o lenguaje.
La estructura secundaria responde al modelo de la doble hélice del ADN propuesto
podría desempeñar esta molécula sus funciones, almacenando y transfiriendo la información genética.
El modelo propone los siguientes aspectos estructurales:
- La existencia de dos cadenas polinucleotídicas dextrógiras arrolladas en forma de hélice alrededor de un mismo eje constituyendo así una doble hélice. Ambas cadenas o hebras son antiparalelas, es decir, sus puentes fosfodiéster 3´- 5´internucleotídicos van en direcciones opuestas, una va en sentido 3′ →5′ y la otra en sentido 5′ →3′ .
- Las bases púricas y pirimidínicas de cada una de las cadenas o hebras están apiladas en el interior de la duplohélice, con sus planos paralelos entre sí, y perpendiculares al eje de la doble hélice. Las bases de una cadena están apareadas mediante puentes de hidrógeno con las bases de la otra cadena. Los pares permisibles son A-T y G-C.
- Las dos cadenas antiparalelas de la duplohélice no son idénticas en secuencia ni en composición. En vez de ello, son complementarias entre sí (siempre que en una cadena hay A, en la otra T, y viceversa).
- Las bases se hallan apiladas a una distancia de 0,34 nm de centro a centro, o lo que es lo mismo, es la distancia que separa cada par de bases. En cada vuelta completa de la duplohélice hay exactamente 10 nucleótidos, lo que corresponde a la distancia secundaria repetida de 3,4 nm. La doble hélice tiene unos 2,0 nm de diámetro.
- Las bases relativamente hidrófobas están situadas en el interior de la hélice y los restos polares del azúcar y grupos fosfato cargados negativamente están en la periferia, expuestos al agua, formando el esqueleto externo de la duplohélice.
- La duplohélice resulta estabilizada no sólo por puentes de hidrógeno entre los pares de bases complementarias, sino también por interacciones electrónicas entre las bases apiladas, así como por acciones hidrófobas recíprocas.
Cuando Watson y Crick propusieron en 1953 la hipótesis de la estructura molecular en doble hélice del ADN, surgieron una hipótesis para explicar cómo se encuentra codificada la información genética. Según ésta, la información genética está contenida en la secuencia de los nucleótidos (A, T, G, C) de la molécula de ADN que determina la secuencia de los aminoácidos en las proteínas.
-Características de los distintos RNAs y su función biológica.
El ARN es un polímero constituido por la unión de ribonucleótidos de adenina, guanina,
citosina y uracilo mediante enlaces fosfodiéster en dirección 5´ → 3´.
Existen varios tipos de ARN, con la misma composición química, pero diferente estructura
y función. Los principales tipos son:
El ARN está compuesto por tres moléculas diferentes:
1) Una pentosa, la β -D-ribofuranosa (ribosa).
2) Una base nitrogenada correspondiente a los dos tipos principales que existen:
- Bases púricas: adenina A) y guanina (G).
- Bases pirimidínicas: citosina (C) y uracilo (U).
3) Una molécula de ácido fosfórico.
Existen tres tipos de ARN diferentes:
- El ARN mensajero (ARNm) que sólo posee estructura primaria.
- El ARN transferente (ARNt) posee estructura primaria y estructura secundaria específica
denominada “hoja de trébol”. Esta última estructura se forma gracias a la presencia de
regiones complementarias que permiten el establecimiento de horquillas intracatenarias
(regiones de doble hélice).
- ARN ribosómico (ARNr) posee estructura terciaria que se produce al unirse diferentes
moléculas de ARNr a proteínas ribosomales, originando las subunidades 30 S y 50 S de los
ribosomas procariontes, y las 40 S y 60.
TIPOS DE ARN FUNCION CELULAR LOCALIZACIÓN
ARN mensajero (ARNm) Su función es copiar la
información genética
contenida en el ADN
(transcripción) y transportarla
hasta el citoplasma, donde
tendrá lugar la síntesis
proteica en los ribosomas.
El ARNm se sintetiza en el
núcleo y transmite la
información en el citoplasma.
ARN transferente (ARNt) El ARNt es el encargado de
transportar los aminoácidos
hasta los ribosomas, para
situarlos sobre el ARNm y
formar la cadena polipeptídica
de acuerdo con el mensaje
genético codificado en el
ARN.
Los distintos tipos de ARNt
se sintetizan en el núcleo y
realizan su función en el
citoplasma.
ARN ribosómico (ARNr) La función del ARNr es
constituir, junto con las
proteínas ribosomales, los
ribosomas y, por lo tanto,
participar en la síntesis
proteica.
Los diferentes tipos de ARNr
se sintetizan en el núcleo y
realizan su función en el
citoplasma.
-Naturaleza de las enzimas, estructura molecular, centro activo y el mecanismo de acción enzimática:
Un enzima es un biocatalizador sintetizado por el propio organismo cuya composición química es total o parcialmente proteica. Los biocatalizadores son sustancias que, sin consumirse en el proceso, intervienen en las reacciones químicas, disminuyendo la energía de activación, por consiguiente, aumentando la velocidad de reacción. Los enzimas actúan de dos maneras:
1.- Fijándose a las sustancias reaccionantes o sustratos, de modo que se debiliten sus enlaces y se favorezca su ruptura.
2.- Atrayendo a su superficie las sustancias reaccionantes, de modo que su encuentro y la reacción se produzcan más fácilmente.
Atendiendo a la naturaleza química de los enzimas, distinguimos dos tipos:
a) Holoproteínas: Su molécula está constituida únicamente por residuos de aminoácidos, es decir, es estrictamente proteica.
b) Heteroproteínas u holoenzimas: en esta caso están formados por dos componentes:
- uno de naturaleza proteica denominado apoenzima.
- otro no proteico denominado cofactor. Si éste está ligado covalentemente a la apoenzima recibe el nombre de grupo prostético. La naturaleza del cofactor puede ser inorgánica (caso de muchos iones metálicos como Fe, Cu, Zn, Mg, etc.), u orgánica, entonces el cofactor recibe el nombre de coenzima.
Las apoenzimas son proteínas globulares constituidas por tres tipos de aminoácidos:
- Aminoácidos estructurales: sin función dinámica.
- Aminoácidos de fijación o de unión al sustrato: sus radicales están encargados de orientar al
sustrato y fijarlo estableciendo enlaces débiles con él.
- Aminoácidos catalizadores: modifican al sustrato y lo transforman en producto.
Los dos últimos tipos de aminoácidos (de fijación y catalíticos) constituyen el centro de reacción del enzima.
En general, el mecanismo de reacción enzima-sustrato puede simbolizarse así:
[E]+ [S]®[ES]®[P]+ [E]
La unión enzima-sustrato se produce mediante fuerzas intermoleculares de carácter débil que tienen lugar en la zona específica del enzima que se denomina centro activo o centro catalítico. Es una zona del apoenzima que posee estructura tridimensional en forma de hueco, generalmente hidrofóbica, donde actúan las cadenas laterales de los aminoácidos de fijación y catalíticos. La fijación del sustrato constituye la primera fase del proceso catalítico de la enzima, en la que intervienen sus aminoácidos de fijación, y en donde radica la especificidad. Luego, una vez fijada la molécula de sustrato, actúan los aminoácidos catalíticos transformándolo en producto. Por tanto, elcentro activo del apoenzima es el responsable directo de la acción catalítica y específica del enzima, aunque el resto de la molécula del enzima (constituida por los aminoácidos estructurales) tenga también importancia en el mantenimiento de todo el conjunto.
-Inhibición enzimática: tipos de inhibidores y modo de acción.
En general, el mecanismo de acción enzimática transcurre siempre con la unión del
sustrato al enzima formándose un complejo intermediario entre ambos, imprescindible
para que la reacción química pueda llevarse a cabo. En general, la reacción enzimática
puede simbolizarse así:
[E]+ [S]→ [ES]→ [P]+ [E]
La inhibición enzimática es un proceso por el cual la actividad de un enzima puede
verse disminuida por moduladores capaces de inhibirla, es decir, de disminuir la
actividad y eficacia de ésta. Estos moduladores son los inhibidores enzimáticos que son
moléculas que modifican la velocidad de reacción de un modo negativo.
La inhibición enzimática es de dos tipos: reversible e irreversible.
- La inhibición reversible del enzima tiene lugar cuando se inactiva temporalmente el
centro activo de ésta, si eliminamos la molécula inhibidor del medio el enzima volverá a
desarrollar un funcionamiento normal.
Existen dos formas de inhibición reversible:
- Competitiva: se debe a la presencia de un inhibidor cuya molécula presenta una
conformación espacial lo suficiente similar al sustrato como fijarse en centro activo de
la enzima. Ambas moléculas, inhibidor y sustrato, compiten por la enzima.
liberándose rápidamente los productos. Al fijarse este inhibidor, la enzima es incapaz de
romperlo y, por tanto, de desembarazarse de él con facilidad.
- No competitiva: es debida a un inhibidor que, o bien actúa sobre el complejo enzimasustrato
haciéndolo fijo, o bien se une a la enzima de tal modo que impide el acceso del
sustrato al centro activo.
Por lo tanto, si se trata de un inhibidor competitivo éste se unirá al centro activo del
enzima, mientras que si es no competitivo el lugar de unión será distinto y por ejemplo
se unirá al complejo enzima-sustrato.
- La inhibición irreversible del enzima tiene lugar cuando ésta se inactiva de modo
permanente y no vuelve a tener un funcionamiento normal, el caso de algunos venenos.
TDC CITOLOGIA
J10B
Las células
eucariotas: Definición,origen,tipos y partes.
1-Definición
de célula eucariota:
Se denominan células
eucariotas a todas las
células que tienen su material hereditario, su información genética,
encerrado dentro de una doble bicapa lipídica: la envoltura
nuclear; la cual delimita un núcleo
celular, al contrario
que las procariotas que carecen de dicha membrana
nuclear, por lo que el material genético se encuentra disperso en ellas (en su citoplasma).
2-Origen de los eucariotas.
El origen de los eucariotas se encuentra en sucesivos
procesos simbiogenéticos (procesos simbióticos que culminan
en la unión de sus simbiontes, estableciéndose una nueva individualidad de los
integrantes) entre diferentes bacterias.
Reuniendo las
investigaciones de Lynn Margulis y otros podemos decir que una célula a la que
llamaremos “urcariota” incorpora diferentes tipos de bacterias que la evolución
“convertirán” en orgánulos como las mitocondrias, cloroplastos, cilios etc.
3- Organismos eucariontes
Los organismos
eucariontes forman
el dominio Eukarya que incluye a los organismos
más conocidos, repartidos en cuatro reinos: Animalia (animales), Plantae (plantas), Fungi(Hongos)
y Protista .
4- Diferencias entre células eucariotas.
Existen diversos tipos de células eucariotas entre las que
destacan las células de animales y plantas. Los hongos y muchos protistas
tienen, sin embargo, algunas diferencias substanciales.
Así pues las
diferencias entre la c. animal y vegetal son:
Las primeras poseen
centríolos, glicocálix, vacuolas numerosas y pequeñas. Las segundas tienen
cloroplastos, vacuolas más grandes (incluso única), pared celular, glioxisomas
y sólo flagelos ciertas células sexuales.
También hay
diferencias a nivel de metabolismo secundario como por ejemplo la síntesis de
esteroides en c. animales o la conversión de lípidos en glúcidos en c.
vegetales.
5- Orgánulos
celulares
Sin distinguir
entre células, que tendrán formas, tamaños y más o menos de uno u otro orgánulo
(así por ejemplo hay células sin núcleo como los hematíes, sin ciertos
orgánulos como mitocondrias en algún protozoo…)
-Orgánulo sin membrana.
a) Citoesqueleto
( incluye los filamentos proteicos )
b) Centrosoma
(incluye centríolos,material pericentriolar y áster)
c) Ribosomas.
-Orgánulos con membrana.
A) Una
membrana:
-
Membrana plasmática.
-
Retículo endoplásmico (liso y rugoso)
-
Aparato de Golgi (incluye vesículas de transición y de
secreción)
-
Lisosomas (primarios, secundarios…)
-
Vacuolas (autofágica, heterofágica…)
-
Plastos ( p.e. amilopastos)
-
Peroxisomas
-
Glioxisomas.
-
B) Doble
membrana:
-Núcleo
-Mitocondrias.
-Cloroplastos.
6- Funciones celulares.
Se refieren y coinciden con las de cualquier ser vivo uni o
pluricelular. Estas son de nutrición, relación y reproducción.
7- Algunas características (diferenciadoras
con las c. procariotas)
a) Tamaño: superior al orden de
micras, normalmente observables al microscopio óptico ,visibles los
“huevos”,ciertas células nerviosa… ( las procariotas micras )
b) Variadas según el reino o
tejido si es el caso ( las p. cocos, bacilos espirilos…)
c) Ribosomas 80 s ( 70s en p. )
d) ADN unido a proteínas.
e) Núcleo.
f) División celular compleja
-mitosis y meiosis.
g) Varios cromosomas.
h) Genes discontinuos (intrones y
exones)
i) Sistema de endomembranas (visto)
j) enzimas respiratorios en
orgánulos (p. en “mesosomas”)
h) Normalmente aerobias.
S10B
-Establecer claramente las diferencias entre el transporte
activo y el transporte pasivo a través de membranas. ¿En qué se diferencian la
difusión simple y la difusión facilitada? (3 puntos)
TRANSPORTE PASIVO
El transporte pasivo es el intercambio simple de moléculas a
través de la membrana plasmática, durante el cual la célula no gasta energía,
debido a que va a favor del gradiente de concentración o a favor de gradiente
de carga eléctrica, es decir, de un lugar donde hay una gran concentración a
uno donde hay menor. El proceso se realiza por difusión(Ósmosis en medio líquido y a través de una membrana semipermeable). En sí,
es el paso de sustancias desde un medio en el que estén a mayor concentración (medio hipertónico) a otro a menor concentración (un medio hipotónico).
Difusión simple: Significa que la molécula puede pasar directamente a través de la membrana. La difusión es siempre a favor de un gradiente de concentración. Esto limita la máxima concentración posible en el interior de la célula (o en el exterior si se trata de un producto de desecho).La efectividad de la difusión está limitada por la velocidad de difusión de la molécula. Por lo tanto si bien la difusión es un mecanismo de transporte suficientemente efectivo para algunas moléculas (por ejemplo el agua), la célula debe utilizar otros mecanismos de transporte para sus necesidades. Difusión simple a través de canales .Se realiza mediante las denominadas proteínas de canal. Así entran iones como el Na.+, K+, Ca2+, Cl-. Las proteínas de canal son proteínas con un orificio o canal interno, cuya apertura está regulada, por ejemplo por ligando, como ocurre con neurotransmisores u hormonas, que se unen a una determinada región, el receptor de la proteína de canal, que sufre una transformación estructural que induce la apertura del canal. Difusión simple a través de la bicapa .Así entran moléculas lipídicas como las hormonas esteroideas, anestésicos como el éter y fármacos liposolubles. Y sustancias apolares como el oxígeno y el nitrógeno atmosférico. Algunas moléculas polares de muy pequeño tamaño, como el agua, el CO2, el etanol y la glicerina, también atraviesan la membrana por difusión simple. La difusión del agua recibe el nombre de ósmosis.
Difusión facilitada: Algunas moléculas son demasiado grandes como para difundir a través de los canales de la membrana y demasiado insolubles en lípidos como para poder difundir a través de la capa de fosfolípidos. Tal es el caso de la glucosa y algunos otros monosacáridos. Estas sustancias, pueden sin embargo cruzar la membrana plasmática mediante el proceso de difusión facilitada, con la ayuda de una proteína transportadora. En el primer paso, la glucosa se une a la proteína transportadora, y esta cambia de forma, permitiendo el paso del azúcar. Tan pronto como la glucosa llega al citoplasma, una enzima que añade un grupo fosfato a un azúcar, (Kinesa) transforma la glucosa en glucosa-6-fosfato. De esta forma, las concentraciones de glucosa en el interior de la célula son siempre muy bajas, y el gradiente de concentración exterior --> interior favorece la difusión de la glucosa. La difusión facilitada es mucho más rápida que la difusión simple y depende:
Difusión simple: Significa que la molécula puede pasar directamente a través de la membrana. La difusión es siempre a favor de un gradiente de concentración. Esto limita la máxima concentración posible en el interior de la célula (o en el exterior si se trata de un producto de desecho).La efectividad de la difusión está limitada por la velocidad de difusión de la molécula. Por lo tanto si bien la difusión es un mecanismo de transporte suficientemente efectivo para algunas moléculas (por ejemplo el agua), la célula debe utilizar otros mecanismos de transporte para sus necesidades. Difusión simple a través de canales .Se realiza mediante las denominadas proteínas de canal. Así entran iones como el Na.+, K+, Ca2+, Cl-. Las proteínas de canal son proteínas con un orificio o canal interno, cuya apertura está regulada, por ejemplo por ligando, como ocurre con neurotransmisores u hormonas, que se unen a una determinada región, el receptor de la proteína de canal, que sufre una transformación estructural que induce la apertura del canal. Difusión simple a través de la bicapa .Así entran moléculas lipídicas como las hormonas esteroideas, anestésicos como el éter y fármacos liposolubles. Y sustancias apolares como el oxígeno y el nitrógeno atmosférico. Algunas moléculas polares de muy pequeño tamaño, como el agua, el CO2, el etanol y la glicerina, también atraviesan la membrana por difusión simple. La difusión del agua recibe el nombre de ósmosis.
Difusión facilitada: Algunas moléculas son demasiado grandes como para difundir a través de los canales de la membrana y demasiado insolubles en lípidos como para poder difundir a través de la capa de fosfolípidos. Tal es el caso de la glucosa y algunos otros monosacáridos. Estas sustancias, pueden sin embargo cruzar la membrana plasmática mediante el proceso de difusión facilitada, con la ayuda de una proteína transportadora. En el primer paso, la glucosa se une a la proteína transportadora, y esta cambia de forma, permitiendo el paso del azúcar. Tan pronto como la glucosa llega al citoplasma, una enzima que añade un grupo fosfato a un azúcar, (Kinesa) transforma la glucosa en glucosa-6-fosfato. De esta forma, las concentraciones de glucosa en el interior de la célula son siempre muy bajas, y el gradiente de concentración exterior --> interior favorece la difusión de la glucosa. La difusión facilitada es mucho más rápida que la difusión simple y depende:
·
Del gradiente de concentración de la sustancia a ambos lados de la
membrana
·
Del número de proteínas transportadoras existentes en la membrana
·
De la rapidez con que estas proteínas hacen su trabajo
Ósmosis :
La ósmosis es un tipo especial de transporte pasivo en el cual sólo las moléculas de agua son transportadas a través de la membrana. El movimiento de agua se realiza desde un punto en que hay mayor concentración a uno de menor para igualar concentraciones. De acuerdo al medio en que se encuentre una célula, la ósmosis varía. La función de la osmosis es mantener hidratada a la membrana celular. Dicho proceso no requiere gasto de energía. En otras palabras la ósmosis u osmosis es un fenómeno consistente en el paso del solvente de una disolución desde una zona de baja concentración de soluto a una de alta concentración del soluto, separadas por una membrana semipermeable.
TRANSPORTE ACTIVO:
El transporte activo requiere un gasto de energía para
transportar la molécula de un lado al otro de la membrana, pero el transporte
activo es el único que puede transportar moléculas contra
un gradiente de concentración, el transporte activo esta limitado
por el número de proteínas transportadoras presentes. Son de interés dos
grandes categorías de transporte activo, primario y secundario(en éste último se aprovecha el paso de iones a favor de gradiente para transportar en contra de gradiente moléculas como glucosa o aminoácidos,puede ser en la misma dirección (simporte) o en direcciones contrarias (antiporte).
Transporte activo( primario): En este caso, la energía derivada del ATP directamente empuja a la sustancia para que cruce la membrana, modificando la forma de las proteínas de transporte (bomba) de la membrana plasmática. El ejemplo más característico es la bomba de Na+/K+, que mantiene una baja concentración de Na+ en el citosol extrayéndolo de la célula en contra de un gradiente de concentración. También mueve los iones K+ desde el exterior hasta el interior de la célula pese a que la concentración intracelular de potasio es superior a la extracelular. Esta bomba debe funcionar constantemente ya que hay pérdidas de K+ y entradas de Na+ por los poros acuosos de la membrana. Esta bomba actúa como una enzima que rompe la molécula de ATP y también se llama bomba Na+/K+-ATPasa. Todas las células poseen cientos de estas bombas por cada μm² de membrana.
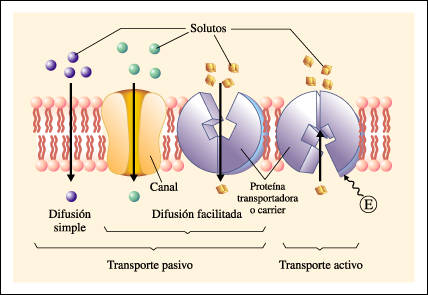
Transporte activo( primario): En este caso, la energía derivada del ATP directamente empuja a la sustancia para que cruce la membrana, modificando la forma de las proteínas de transporte (bomba) de la membrana plasmática. El ejemplo más característico es la bomba de Na+/K+, que mantiene una baja concentración de Na+ en el citosol extrayéndolo de la célula en contra de un gradiente de concentración. También mueve los iones K+ desde el exterior hasta el interior de la célula pese a que la concentración intracelular de potasio es superior a la extracelular. Esta bomba debe funcionar constantemente ya que hay pérdidas de K+ y entradas de Na+ por los poros acuosos de la membrana. Esta bomba actúa como una enzima que rompe la molécula de ATP y también se llama bomba Na+/K+-ATPasa. Todas las células poseen cientos de estas bombas por cada μm² de membrana.
-Estructura de las células procariotas.
Existen dos grandes tipos celulares: las procariotas (que comprenden las células de arqueas y bacterias) y las eucariotas.
Las células procariotas son pequeñas y menos complejas que las eucariotas. Contienen ribosomas(70s) pero carecen de sistemas de endomembranas (esto es, orgánulos delimitados por membranas biológicas, como puede ser el núcleo celular). Por ello poseen el material genético en el citosol. Sin embargo, existen excepciones: algunas bacterias fotosintéticas poseen sistemas de membranas internos. Por lo general podría decirse que los procariotas carecen de citoesqueleto. Sin embargo se ha observado que algunas bacterias, como Bacillus subtilis.
En el citoplasma se pueden apreciar plásmidos(elemento facultativo), pequeñas moléculas circulares de ADN que coexisten con el nucleoide y que contienen genes: son comúnmente usados por las bacterias en la parasexualidad,resistencia a los antibióticos...
Poseen una membrana celular (Contiene las enzimas de la cadena respiratoria, también puede poseer los pigmentos fotosintéticos)compuesta de lípidos, en forma de una bicapa y sobre ella se encuentra una cubierta en la que existe un polisacárido complejo denominado peptidoglicano; dependiendo de su estructura y subsecuente su respuesta a la tinción de Gram, se clasifica a las bacterias en Gram positivas y Gram negativas. El espacio comprendido entre la membrana celular y la pared celular (o la membrana externa, si ésta existe) se denomina espacio periplásmico. Las arqueas poseen una pared celular de composición característica, por ejemplo, no contienen peptidoglicano (mureína) como en las bacterias.
Algunas bacterias presentan una cápsula. Otras son capaces de generar endosporas (estadios latentes capaces de resistir condiciones extremas) en algún momento de su ciclo vital. Entre las formaciones exteriores propias de la célula bacteriana destacan los flagelos (de estructura completamente distinta a la de los flagelos eucariotas) y los pili (estructuras de adherencia y relacionadas con la parasexualidad).
-Morfología de las células procariotas y organización de su material genético.
a-Las células procariotas son pequeñas y menos complejas que las eucariotas. Por lo general podría decirse que los procariotas carecen de citoesqueleto. (Sin embargo se ha observado en algunas bacterias, como Bacillus subtilis, ). El tamaño es un parámetro que está determinado genéticamente, pero los valores concretos para cada raza o cepa de bacterias vienen influidos por una serie de condiciones ambientales (nutrientes, sales, temperatura, tensión superficial, etc).
-Tipos según su morfología:
Coco es un tipo morfológico de bacteria. Tiene forma más o menos esférica (ninguna de sus dimensiones predomina claramente sobre las otras).
Los bacilos son bacterias que tienen forma de bastón, cuando se observan al microscopio. Los bacilos se suelen dividir en:
Bacilos Gram positivos: fijan el violeta de genciana (tinción de Gram) en la pared celular porque carecen de capa de lipopolisacáridos.
Bacilos Gram negativos: no fijan el violeta de genciana porque poseen la capa de lipopolisacárido.
Varias de las especies de Vibrio son patógenas, provocando enfermedades del tracto digestivo, en especial V. cholerae, el agente que provoca el cólera.
Los espirilos son bacterias flageladas de forma helicoidal o de espiral. Se desplazan en medios viscosos avanzando en tornillo. Su diámetro es muy pequeño, lo que hace que puedan atravesar las mucosas; por ejemplo Treponema pallidum que produce la sífilis en el hombre.
En algunas especies las células hijas pueden permanecer unidas entre sí (al menos durante un cierto tiempo tras la división de la que proceden) debido a que el tabique sea incompleto o a la existencia de capas mucosas que retienen juntos los productos de la división.
Si la tendencia a permanecer unidas es baja, tendremos agrupaciones de d células:-diplococos-diplobacilos-estreptococos (cadenetas arrosariadas de cocos) y estreptobacilos (cadenetas de bacilos).-tétradas (4 céls. en un plano) o múltiplos;-sarcinas (paquetes cúbicos);-estafilococos (racimos irregulares).-bacilos en empalizada o en paquetes de cigarrillos (debido a giros de 180o)-dos bacilos en ángulo (en forma de letra V o L)-varios bacilos formando “letras chinas”.
Por otra parte las bacterias pueden o no tener cápsula (estructura superficial que presentan muchas bacterias consistente en acumulación de material mucoso o viscoso),condicionando ésta la morfología.
También tenemos fenómenos de multicelularidad p.ej. mixobacterias,filamentos de cianobacterias

b-Material genético de las bacterias.
Nucleoide (que significa Similar al núcleo y también se conoce como Región nuclear o Cuerpo nuclear) es la región que contiene el ADN en el citoplasma de las células procariotas. Esta región es de forma irregular.
En las células procariotas, el ADN es una molécula única, generalmente circular y de doble filamento y superenrrollado, que se encuentra ubicada en un sector de la célula que se conoce con el nombre de nucleoide, que no implica la presencia de membrana nuclear. Dentro del nucleoide pueden existir varias copias de la molécula de ADN.
Este sistema para guardar la información genética contrasta con el sistema existente en células eucariotas, donde el ADN se guarda dentro de un orgánulo con membrana propia llamado núcleo.
el nucleoide está compuesto fundamentalmente por ADN (60%), con pequeñas proporciones de ARN y proteínas. Estos dos últimos componentes actúan como ARN mensajero y como proteínas reguladoras del genoma.
Plasmidos
 Los plásmidos, vectores o también llamados plasmidios, son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Las moléculas de ADN plásmidico, adoptan una conformación tipo doble hélice al igual que el ADN de los cromosomas, aunque, por definición, se encuentran fuera de los mismos. Se han encontrado plásmidos en casi todas las bacterias. En la mayoría de los casos se considera genético dispensable. Sin embargo, posee información genética importante para las bacterias. Por ejemplo,los genes que codifican para las proteínas que las hace resistentes a los antibióticos están, frecuentemente, en los plásmidos.
Los plásmidos, vectores o también llamados plasmidios, son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Las moléculas de ADN plásmidico, adoptan una conformación tipo doble hélice al igual que el ADN de los cromosomas, aunque, por definición, se encuentran fuera de los mismos. Se han encontrado plásmidos en casi todas las bacterias. En la mayoría de los casos se considera genético dispensable. Sin embargo, posee información genética importante para las bacterias. Por ejemplo,los genes que codifican para las proteínas que las hace resistentes a los antibióticos están, frecuentemente, en los plásmidos.Hay algunos plásmidos integrativos, es decir, que tienen la capacidad de insertarse en el cromosoma bacteriano. Estos rompen momentaneamente el cromosoma y se sitúan en su interior, con lo cual, automáticamente la maquinaria celular también reproduce el plásmido. Cuando ese plásmido se ha insertado se les da el nombre de episoma.
Otra forma de clasificar plásmidos es por su función. Hay 5 clases principales:
Plásmidos de fertilidad: los cuales contienen tra-genes, son capaces de conjugarse.
Plásmidos de resistencia: los cuales contienen genes que pueden constituir resistencia contra antibióticos o venenos. Históricamente conocidos como Factores R, antes de que se entendiera la naturaleza de los plásmidos.
Col-plásmidos ,Plásmidos degradativos y Plásmidos virulentos
6-La célula procariota y sus características diferenciales con respecto a la célula eucariota.
Ver tabla comparación libro.
-Membrana plasmática. Composición química y estructura. Modelos de membrana. Propiedades y funciones.
Del análisis bioquímico de las membranas plasmáticas aisladas se deduce que estas están
compuestas por lípidos, proteínas y en menor proporción glúcidos:
- Las membranas biológicas de las células eucariotas están compuestas por tres tipos de
lípidos: fosfolípidos, glucolípidos y colesterol. Éstos se encuentran formando una bicapa
lipídica que aporta la estructura básica a la membrana y actúa de barrera relativamente
impermeable al flujo de la mayoría de las moléculas hidrosolubles.
- Las proteínas median las diversas funciones de la membrana. La mayoría poseen estructura
globular y según su posición en la membrana se clasifican en dos tipos: proteínas integrales
transmembrana o intrínsecas, que están intercaladas o embebidas en la bicapa lipídica, y las
proteínas periféricas o extrínsecas , que no atraviesan la membrana y generalmente se
encuentran situadas en el exterior. Estas últimas se encuentran unidas a los lípidos de la
bicapa mediante enlaces covalentes, o las proteínas transmembrana mediante puentes de
hidrógeno.
- Los glúcidos están representados por oligosacáridos que forman el llamado glicocálix en la
superficie externa de la membrana. La gran mayoría están unidos covalentemente a los lípidos
o a las proteínas formando glucolípidos y glucoproteínas respectivamente.
En la actualidad el modelo de estructura de la membrana plasmática más aceptado es el
“modelo del mosaico fluido” propuesto por Singer y Nicolson en 1972. Según este modelo
las membranas poseen lípidos, proteínas y oligosacáridos que se disponen formando una
configuración de baja energía libre. Se dice que la membrana es un mosaico fluido, porque
según este modelo las membranas biológicas son estructuras casi fluidas, dinámicas, en las
que tanto los lípidos como las proteínas pueden realizar constantes movimientos de traslación
lateral, de la bicapa lípidica, aunque los movimientos lípidos de una capa a otra o en “flipflop”
son raros. Además, las membranas son estructuras asimétricas, puesto que la
composición lipídica y proteica de sus dos caras es diferente de modo que refleja las
diferentes funciones realizadas por las dos superficies.
Los lípidos se encuentran formando una bicapa lipídica que aporta la estructura básica a la
membrana y actúa de barrera relativamente impermeable al flujo de la mayoría de las
moléculas hidrosolubles. Las moléculas están orientadas de forma que los grupos polares se
dirigen hacia la fase acuosa, es decir, los de la capa exterior de la membrana hacia el medio
extracelular y los de la capa interior hacia el citoplasma. Además de sus propiedades de autoensamblaje
y auto-sellado, las bicapas lipídicas tienen otra característica que las convierte en
una estructura ideal para las membranas celulares, esta característica es su fluidez, que
permite que las moléculas lipídicas puedan desplazarse libremente por la membrana, en
movimientos laterales o de rotación sobre sí mismos.
El colesterol el encargado de regular la fluidez de la bicapa, ya que éste interfiere con las
cadenas hidrocarbonadas de los ácidos grasos y les confiere rigidez, a la vez que impide que
las cadenas se junten y agreguen.
La membrana plasmática desarrolla funciones muy importantes para la célula, entre ellas
citamos las siguientes:
1.- Sirve de frontera física entre el medio intra y extracelular.
2.- Mantiene la permeabilidad selectiva, mediante el control del intercambio de sustancias
entre el medio y la célula de modo controlado y selectivo.
La mayor parte de las funciones que desempeña la membrana plasmática son responsabilidad
directa de las proteínas que la componen. Éstas median diversas funciones: algunas sirven
para el transporte de moléculas específicas hacia el exterior e interior de la célula; otras son
enzimas que catalizan reacciones asociadas a la membrana, otras actúan de eslabones entre el
citoesqueleto de la célula y la matriz extracelular, o bien actúan como receptores que reciben
y traducen las señales químicas procedentes del entorno de la célula.
Otras: ver libro.
-El núcleo interfásico: describa su morfología al microscopio electrónico, la constitución
química y la estructura de cada uno de sus componentes, indicando su función.
El núcleo es el componente celular donde se encuentra el material genético y que dirige toda
la vida celular. Cuando se trata de un núcleo en estado interfásico presenta una estructura,
morfología y composición típicas ya que durante la división celular la membrana desaparece
y sus componentes se reparten entre lo que serán las dos células hijas.
En un núcleo interfásico distinguimos los siguientes componentes:
- La membrana nuclear representa una compleja frontera entre el núcleo y el citoplasma de
una célula eucariota. Al microscopio electrónico se observa que es doble con un espacio
intermembranoso.
- La membrana nuclear externa presenta sobre su cara externa ribosomas
adosados. Esta membrana suele estar unida al retículo endoplásmico, ya sea liso o
rugoso.
- El espacio perinuclear o intermembranoso está comprendido entre las dos
membranas.
- La membrana nuclear interna presenta, asociado a ella, en la cara
nucleoplásmica, un material electrodenso de naturaleza fibrilar denominado
lámina fibrosa. A esta estructura se le atribuyen funciones de servir de anclaje a la
cromatina y regular el crecimiento de la envoltura nuclear.
- En el interior del núcleo encontramos una sustancia filamentosa que corresponde a la
cromatina que se encuentra inmersa en el denominado nucleoplasma que consiste en una
matriz semifluida. Además, dentro del núcleo aparecen uno o dos cuerpos esféricos que son
los nucléolos, responsables de la síntesis del ARNr, de su procesamiento y ensamblaje con
proteínas, dando lugar a las subunidades ribosómicas que posteriormente salen al citoplasma
para constituir los ribosomas.
En todos los núcleos, las dos membranas que forman la envoltura nuclear se fusionan en
algunos lugares, dando origen a unas perforaciones circulares denominadas poros nucleares.
Se trata de estructuras dinámicas, capaces de formarse y desaparecer, dependiendo del estado
funcional de la célula. Son canales acuosos que regulan los intercambios de moléculas entre el
núcleo y el citosol.
Cuando se utilizan el microscopio electrónico se observa que los poros no son simples
orificios, sino estructuras complejas que se denominan complejo del poro. Constan de:
- Un anillo o estructura cilíndrica que se proyecta tanto hacia la cara citoplasmática como a la
nucleoplasmática, y que está constituida por ocho partículas proteicas dispuestas en
octógonos. Asociado a estas partículas se encuentra un material denso, el diafragma, que
disminuye la luz del poro.
- En ocasiones se ha identificado en el centro del poro un gránulo central, que en realidad se
corresponde a ribosomas recién formados o sustancias que van de un lado a otro de la
membrana nuclear.
-El núcleo: la envoltura nuclear, el nucleoplasma, nucleolos, cromatina.
El núcleo es el componente celular característico de las células eucariotas donde se
encuentra el material genético que dirige la vida celular. Cuando se trata de un núcleo
en estado interfásico presenta una estructura, morfología y composición típicas ya que
durante la división celular la membrana desaparece y sus componentes se reparten entre
lo que serán las dos células hijas.
En un núcleo interfásico distinguimos los siguientes componentes:
- La envoltura nuclear representa una compleja frontera entre el núcleo y el citoplasma
de una célula eucariota. Al microscopio electrónico se observa que es doble con un
espacio intermembranoso.
- La membrana nuclear externa presenta sobre su cara externa ribosomas
adosados. Esta membrana suele estar unida al retículo endoplásmico, ya sea liso
o rugoso.
- El espacio perinuclear o intermembranoso está comprendido entre las dos
membranas.
- La membrana nuclear interna presenta, asociado a ella, en la cara
nucleoplásmica, un material electrodenso de naturaleza fibrilar denominado
lámina fibrosa. A esta estructura se le atribuyen funciones de servir de anclaje a
la cromatina y regular el crecimiento de la envoltura nuclear.
En todos los núcleos, las dos membranas que forman la envoltura nuclear se fusionan en
algunos lugares, dando origen a unas perforaciones circulares denominadas poros
nucleares. Se trata de estructuras dinámicas, capaces de formarse y desaparecer,
dependiendo del estado funcional de la célula. Son canales acuosos que regulan los
intercambios de moléculas entre el núcleo y el citosol.
- En el interior del núcleo encontramos una sustancia filamentosa que corresponde a la
cromatina. Se trata de una estructura compacta y empaquetada formada por ADN
asociado a proteínas.
- La matiz semifluida situada en el interior del núcleo que contiene tanto el material
cromatínico como el no cromatínico se denomina nucleoplasma.
- Además, dentro del núcleo aparecen uno o dos cuerpos esféricos que son los
nucléolos, responsables de la síntesis del ARNr, de su procesamiento y ensamblaje con
proteínas, dando lugar a las subunidades ribosómicas que posteriormente salen al
citoplasma para constituir los ribosomas.
Cuando el núcleo comienza a dividirse, la cromatina se condensa para formar los
cromosomas. Los nucléolos también dejan de ser visibles.
Si observamos el núcleo de una célula eucariota cuando se encuentra dividiéndose sólo
observamos los cromosomas.
-Composición, estructura y función de los centríolos.
El centriolo es un orgánulo que generalmente se sitúa cerca del núcleo o en el lado
cóncavo del aparato de Golgi de y que solamente está presente en células eucariotas
animales.
Hay un par de centriolos por célula y se disponen perpendiculares entre sí,
constituyendo el diplosoma. Cada centriolo está rodeado por una corona de material
denso que forma acúmulos denominados satélites. El conjunto de diplosoma y satélites
constituyen el centrosoma o centro organizador de microtúbulos.
El centriolo está constituido por 9 tripletes de microtúbulos constituyendo en conjunto
un cilindro. Los microtúbulos del triplete se denominan, de dentro a fuera A, B, y C, y
están, en parte, fusionados. Por ello, sólo el microtúbulo A posee sección redonda,
mientras que los otros la tienen semilunar. Todos los microtúbulos A poseen dos brazos
mal delimitados, de forma de uno de ellos se dirige al microtúbulo C del triplete
adyacente y el otro al centro del cilindro centriolar.
Los microtúbulos son los principales componentes del citoesqueleto de las células
eucarióticas. Son de 250 Å de diámetro y están formados por proteínas globulares,
denominadas tubulinas, que se disponen helicoidalmente de forma que cada vuelta hay
13 unidades que dejan un hueco central. Aparecen libres en el citoplasma, aunque la
mayoría se disponen radialmente al centrosoma, que es una región próxima al núcleo
celular, considerada como el centro organizador de microtúbulos.
Los centriolos se duplican durante la división celular, siendo este proceso el que genera
los polos del huso acromático que permiten la división del núcleo celular en dos,
durante la mitosis. También intervienen en la formación del huso acromático, así como
en la formación de cilios y flagelos y en el movimiento de éstos.
-El aparato de Golgi: estructura y función.
El aparato de Golgi es un orgánulo membranoso constituido por conjunto de sáculos
discoidales y aplanados, delimitados por una unidad de membrana, ligeramente dilatados en
sus extremos de donde parecen desprenderse vesículas. Estos sáculos se agrupan en pilas de 5
a 10 unidades que se denominan dictiosomas, los cuales suelen presentar una superficie
cóncava y otra convexa. Las cavidades están delimitadas por una membrana unitaria y están
llenas de fluido. Los diferentes dictiosomas están conectados entre sí. El aparato de Golgi
suele encontrarse rodeando al núcleo o al centrosoma.
El aparato de Golgi guarda una estrecha relación entre su estructura y su función, y, a su vez,
se relaciona con el retículo endoplásmico, ya que se forma a sus expensas, y sus funciones son
complementarias, retículo y aparato de Golgi forman el denominado complejo GERL. Las dos
superficies o caras del aparato de Golgi delimitan tres espacios en el mismo:
- Cara cis (externa o de formación): es la cara cóncava de los sáculos que está rodeada por
cisternas de retículo endoplásmico que, por gemación, desprenden vesículas. Estas vesículas
se denominan vesículas de transición y están cargadas de productos almacenadas en el
retículo endoplásmico. Varias de ellas se fusionan con los sáculos del aparato de Golgi,
constituyendo en primer espacio del mismo. Desde aquí se van desprendiendo nuevas
vesículas que van circulando hacia la cara convexa dando lugar al tercer compartimento del
aparato de Golgi.
- Cara trans (interna o de maduración): Al llegar a la cara convexa del dictiosoma se
fragmentan en vesículas de secreción. Varias de esas vesículas pueden fusionarse y formar
gránulos de secreción. Estos últimos pueden permanecer en el citoplasma o ir al espacio
extracelular por exocitosis.
El aparato de Golgi interviene en la glicoxilación (unión de glúcidos) a proteínas y lípidos de
membrana procedentes del retículo endoplásmico, que van a ser productos de secreción
celular. Además, interviene en la producción de membranas celulares y en la formación de
lisosomas.
Otras: ver libro.
-Pared celular vegetal: composición, estructura y función.
La pared celular o membrana de secreción vegetal es una forma especializada de
matriz extracelular, que se encuentra adosada a la membrana plasmática de las células
vegetales. Aunque en la superficie de las células animales también existen componentes
de la matriz extracelular, adheridos al glucocálix, la pared celular de los vegetales está
más organizada y es más rígida. Esta rigidez, unida al grosor de la pared, permite a los
vegetales prescindir de un esqueleto.
Al estudiar su estructura se observan dos componentes claramente diferenciados: las
moléculas fibrilares de celulosa, cuya agregación da origen a las fibras de celulosa;
están formadas por unas 1.500 microfibrillas y son visibles con la lupa; y el cemento
que une las fibras de celulosa, formado por pectinas, hemicelulosa, agua y sales
minerales.
Aunque la composición varía mucho de unas células a otras, el principio arquitectónico
en el que se basa es constante: fibras largas y muy resistentes unidas por una matriz de
proteínas y polisacáridos.
El modo en que se organizan estos componentes para formar la pared, se comprende
mejor si seguimos la evolución de una célula que acaba de dividirse, dando origen a dos
céluas hijas que, como ocurre en los vegetales, se mantienen juntas.
La primera capa que se forma a partir de la membrana plasmática es la lámina media,
común a las dos células y formada principalmente por pectinas.
Entre la lámina media y la membrana plasmática depositan hasta 3 capas, dando lugar a
la pared primaria. En ella, las moléculas de celulosa se disponen en red y abunda el
cemento.
Cuando la célula deja de crecer puede conservar la pared primaria, a veces
engrosándola, o deposita nuevas capas de distinta composición, originando así la pared
secundaria.
Normalemente se trata de 3 capas pero, en ocasiones, llegan a ser 20. En ellas
predomina la celulosa sobre el cemento. Las moléculas de células están ordenas y
dispuestas paralelamente, lo que confiere gran resitencia a la pared, aunque la célual
pierde la capacidad de estirarse.
TDC METABOLISMO
1-El papel del ATP en los seres vivos.
El ATP es un nucleótido(derivado) no nucleico, es decir, no forma parte de los ácidos
nucléicos. Químicamente está compuesto por diferentes unidades estructurales que son:
- Una base nitrogenada púrica, la adenina.
- Una pentosa, la ribosa.
- Tres moléculas de ácido ortofosfórico.
El ATP (adenosín trifosofato) es nucleótido complejo de gran importancia biológica, no sólo
como coenzima, sino también por la energía bioquímica que es capaz de almacenar en
sus dos enlaces esterfosfóricos. Al romperse estos enlaces mediante hidrólisis, liberan
cada uno 7,3 Kcal/mol (proceso exergónico).
Por el contrario, la fosforilación del ADP a ATP es un proceso endergónico, no
espontáneo , que requiere un aporte energético.
Las reacciones metabólicas están acopladas energéticamente a través del ATP. En el
metabolismo celular tienen lugar reacciones que liberan energía y otras que la consumen
(en el catabolismo se libera energía y en el anabolismo se consume). Estos procesos
energéticos no tienen por qué ocurrir al mismo tiempo ni en el mismo lugar en la célula.
Por lo tanto, debe existir un mecanismo que almacene y transporte esa energía desde los
lugares donde se libera a los lugares donde se consume. Este mecanismo se base en la
formación y posterior ruptura de enlaces químicos que almacenan y liberan gran
cantidad de energía. Estos enlaces se denominan enlaces de alta energía.
La síntesis del ATP tiene lugar en la célula mediante 3 mecanismos diferentes:
1) Fosforilación a nivel de sustrato: el ATP se produce a partir de ADP al que se une
un fosfato transferido desde otra molécula al que estaba unido.
2) Fosforilación en el transporte electrónico: el transporte de electrones, a través de
proteínas ubicadas en la membrana de mitocondrias y cloroplastos, libera energía que es
utilizada por el enzima ATP-asa para fosforilar el ADP a ATP. Se denomina
fotofosforilación si se produce en el cloroplasto y fosforilación oxidativa si tiene lugar
en la mitocondria.
3)Fotofosrilación: similar a 1) pero en los tilacoides de los cloroplastos de las c. vegetales.
Algunos procesos celulares vitales que requieren la hidrólisis del ATP para su realización
son:
- Transporte activo de sustancias a través de la membrana plasmática.
- Traducción del mensaje genético.
- Movimiento celular.
2-La cadena respiratoria y la fosforilación oxidativa: descripción global de los procesos, y su localización celular.
En conjunto, la cadena de transporte de electrones y la quimiosmosis constituyen la fosforilación oxidativa. Los pasos clave de este proceso, incluyen:
- Entrega de electrones por NADH y FADHstart subscript, 2, end subscript. Los acarreadores de electrones (NADH y FADHstart subscript, 2, end subscript) reducidos en otros pasos de la respiración celular transfieren sus electrones a las moléculas cercanas al inicio de la cadena de transporte. En el proceso se convierten en NADstart superscript, plus, end superscript y FAD, que pueden ser reutilizados en otros pasos de la respiración celular.
- Transferencia de electrones y bombeo de protones. Conforme se mueven los electrones en la cadena, se desplazan de un nivel de energía más alto a uno más bajo, lo que libera energía. Parte de esta energía se utiliza para bombear iones de Hstart superscript, plus, end superscript, lo que los desplaza fuera desde la matriz hacia el espacio intermembranal. Este bombeo establece un gradiente electroquímico.
- Separación de oxígeno molecular para formar agua. Al final de la cadena de transporte de electrones, los electrones se transfieren a una molécula de oxígeno, la cual se rompe a la mitad y recolecta Hstart superscript, plus, end superscript para formar agua.
- Síntesis de ATP impulsada por un gradiente. Cuando fluyen por el gradiente de regreso hacia la matriz, los iones de Hstart superscript, plus, end superscript pasan a través de una enzima llamada ATP sintasa, la cual aprovecha el flujo de protones para sintetizar ATP.


3-Fotosíntesis. Estructuras subcelulares que intervienen en el proceso, Dibújalas y
enumera sus partes. Fase luminosa y fase oscura.
La fotosíntesis es un proceso anabólico y autotrófico primordial, del que depende la vida
sobre la Tierra. Consiste en la conversión por los organismos fotosintéticos de la energía
luminosa procedente del Sol en energía eléctrica y después en energía química. Esta energía
será utilizada para formar materia orgánica propia o biomasa (glúcidos) a partir de moléculas
inorgánicas, como agua, CO2 y sales minerales. El O2 molecula r, resultante de la ruptura de
moléculas de agua que intervienen en el proceso, se desprende como producto de desecho.
Los cloroplastos son unos orgánulos citoplasmáticos cuya función principal es realizar la
fotosíntesis.. Se encuentran rodeados por dos membranas entre las que existe un espacio
intermembranoso. La membrana plastidial externa es lisa, mientras que la membrana
plastidial interna posee invaginaciones paralelas al eje longitudinal del cloroplasto que dan
lugar a la membrana tilacoidal (laminillas, lamelas o tilacoides). La membrana interna encierra un espacio llamado estroma. La membrana tilacoidal se organiza formando unas
vesículas discoidales y aplanadas que se superponen como pilas de monedas llamadas grana.

La fotosíntesis tiene lugar en dos etapas conocidas como fase luminosa y fase oscura.
La fase luminosa de la fotosíntesis tiene lugar en presencia de luz y comprende un conjunto
de reacciones que tienen lugar en las membranas de los tilacoides. En ella, los electrones
liberados tras la incidencia de los fotones sobre los fotosistemas se utilizan para reducir el
NADP+ a NADPH. A través de la cadena de transporte electrónico, la energía de los
electrones se utiliza para la síntesis de ATP. Por tanto, durante esta fase tienen lugar dos
procesos muy importantes: la fotólisis del agua por la que se obtiene poder reductor en forma
de coenzimas reducidas (NADPH), y la fotosfosforilación que produce ATP. El producto de
desecho de esta fase es el oxígeno molecular.
En general, las reacciones luminosas se desarrollan de la siguiente manera:
- La fotólisis del agua produce gracias a la luz H+ que reducirán el CO2 a materia orgánica en
la fase oscura.
H2O ¾l¾uz®½ O2 + 2H+ + 2e-
Esta reacción también tiene lugar en las membranas de los tilacoides, como todos los procesos
que tienen lugar durante la fase luminosa de la fotosíntesis.
- Absorción o captación de la luz solar: es llevada a cabo por los pigmentos fotosintéticos.
Éstos son las clorofilas y los carotenoides. Estos pigmentos junto a proteínas específicas se
encuentran agrupados formando los llamados fotosistemas, que aparecen ubicados en las
membranas tilacoidales de los cloroplastos. Todos los pigmentos del fotosistema son capaces
de absorber luz, pero sólo uno es capaz de convertir la energía luminosa en eléctrica, es el
denominado centro de reacción, que está formado por una molécula de clorofila a y una
proteína específica. El resto de los pigmentos se denominan colectores o antena del
fotosistema y su función es transformar la energía del fotón en la que es capaz de absorber el
centro de reacción.
En la absorción de la luz intervienen dos fotosistemas:
- Fotosistema I (FSI). Capaz de absorber luz de l <700 nm.
- Fotosistema II (FSII): capaz de absorber luz de l<680 nm.
Si un fotón choca con un electrón de un átomo perteneciente a una molécula de pigmento
fotosintético, este electrón capta la energía del fotón y salta a órbitas más alejadas del núcleo,
pudiendo llegar a perderse dejando ionizado al átomo. La molécula que contiene este átomo
queda asimismo oxidada y busca con avidez electrones, que le son proporcionados por el agua
mediante la fotólisis de esta molécula. Por lo tanto, la absorción de la luz por el centro de
reacción hace que la clorofila existente en éste libere un electrón, que viajará a lo largo de una
cadena de transporte de electrones hasta alcanzar el NADP+. Entonces, el centro de reacción
queda ionizado, por lo que se necesita un electrón que será aportado por el H2O mediante
fotólisis de ésta.
- Transporte o flujo electrónico fotosintético: Los electrones arrancados del centro de reacción
cargados con la energía del fotón, son transportados por un conjunto de proteínas
transportadoras, situadas en la membrana tilacoidal, hasta la coenzima NADP+, que se reduce
a NADPH.
- Fotofosforilación: es la formación de ATP debida a la luz. Para que tenga lugar la fase
oscura de la fotosíntesis se necesita NADPH y ATP. Según la “hipótesis quimiosmótica” de
Mitchell, la energía liberada en le transporte de electrones se utiliza para bombear protones,
en contra de un gradiente, desde el estroma la espacio intratilacoidal. Estos protones regresan
al estroma a favor de gradiente a través del complejo enzimático denominado ATP-asa, que
utilizará la energía liberada en el transporte para fosforilar el ADP y transformarlo en ATP.
En la fase oscura de la fotosíntesis o fase de biosíntesis, el ATP y el NADPH obtenidos en
la fase luminosa son utilizados para transformar los compuestos inorgánicos en orgánicos, es
decir, para reducir el CO2 y así sintetizar glúcidos sencillos.
El ciclo de Calvin es la ruta metabólica mayoritaria por la que los organismos fotosintéticos
fijan el CO2, obteniendo así el carbono necesario para la construcción de sus biomoléculas
orgánicas. El ciclo de Calvin consta de varias fases:
1) Fase carboxilativa: El CO2 se incorpora a una molécula de 5 átomos de carbono, la
ribulosa-1,5-difosfato, para producir dos moléculas de 3-fosfoglicerato, es una reacción
catalizada por el enzima ribulosa-1,5-difosfato-carboxilasa-oxigenasa (RUBISCO), que es la
enzima más abundante del planeta.
2) Fase reductora: en esta fase se reduce el CO2 incorporado en la fase carboxilativa en el 3-
fosfoglicerato en forma de grupo carboxilo, mediante el ATP y el NADPH producidos en la
fase luminosa.
3) Fase regenerativa: en esta fase parte del gliceraldehído-3-fosfato obtenido en la fase
reductiva es convertido en glucosa-6-fosfato, y parte en ribulosa-1,5-difosfato para cerrar de
nuevo el ciclo.
El balance del ciclo de Calvin es:
- 6 CO2 + 18 ATP + 12 NADPH + 12 H+ + 12 H2O ® Glucosa + 18 ADP + 18 Pi + 12 NADP+
Este alto consumo de ATP refleja el hecho de que el CO2 es la forma más altamente oxidada
de la cual se puede obtener carbono para construir los esqueletos carbonados de las moléculas
orgánicas.
4-Fase luminosa de la Fotosíntesis.
La reacción o ecuación global de la fotosíntesis en realidad, se lleva a cabo en dos
etapas o fases distintas: fase luminosa y fase oscura.
Se distinguen dos fases luminosas distintas atendiendo a la existencia de dos tipos de
transporte electrónico diferentes, uno es cíclico y el otro es acíclico. Según funcione uno
u otro transporte electrónico se habla de fase luminosa cíclica y fase luminosa acíclica
de la fotosíntesis.
En la fase luminosa de la fotosíntesis acíclica participan los dos fotosistemas (FSI y
FSII) en el transporte electrónico y se produce ATP y poder reductor (NADPH).
La absorción o captación de la luz solar es llevada a cabo por los pigmentos
fotosintéticos. Éstos son las clorofilas, la xantofila y los carotenoides. Estos pigmentos
junto a proteínas específicas se encuentran agrupados formando los llamados
fotosistemas, que aparecen ubicados en las membranas tilacoidales de los cloroplastos.
Todos los pigmentos del fotosistema son capaces de absorber luz, pero sólo uno es
capaz de convertir la energía luminosa en eléctrica, es el denominado centro de
reacción, que está formado por una molécula de clorofila a y una proteína específica. El
resto son los pigmentos antena o complejo antena del fotosistema y su función es
transformar la energía del fotón en la que es capaz de absorber el centro de reacción.
La fotofosforilación es la formación de ATP debida a la luz. Para que tenga
lugar la fase oscura de la fotosíntesis se necesita NADPH y ATP. Según la “hipótesis
quimiosmótica” de Mitchell, la energía liberada en el transporte de electrones desde el
agua hasta el NADP+ se utiliza para bombear protones, en contra de un gradiente, desde
el estroma la espacio intratilacoidal. Estos protones regresan al estroma a favor de
gradiente a través del complejo enzimático denominado ATP-asa, que utilizará la
energía liberada en el transporte para fosforilar el ADP y transformarlo en ATP.
La fotólisis del agua que tiene lugar en la fase luminosa produce H+ que reducirán el
CO2 a materia orgánica en la fase oscura.
H2O ¾l¾uz®½ O2 + 2H+ + 2e-
Esta reacción también tiene lugar en las membranas de los tilacoides, como todos los
procesos que tienen lugar durante la fase luminosa de la fotosíntesis.
5-El ciclo de Calvin o fase “oscura” de la fotosíntesis: localización e interpretación global del proceso.
Durante la fase luminosa o fotoquímica de la fotosíntesis, la energía lumínica ha sido almacenada en moléculas orgánicas sencillas e inestables, que van a aportar energía para realizar el proceso (ATP) y poder reductor, es decir, la capacidad de donar electrones (reducir) a otra molécula (nicotín-amida dinucleótido fosfato o NADPH+H+). En general, los compuestos bioquímicos más reducidos (simplificando la cuestión: los que tienen más electrones) almacenan más energía que los oxidados (con menos electrones, también simplificando) y son, por tanto, capaces de generar más trabajo (por ejemplo, aportar la energía necesaria para el movimiento muscular). En el ciclo de Calvin se integran y convierten moléculas inorgánicas de dióxido de carbono en moléculas orgánicas sencillas a partir de las cuales se formará el resto de los compuestos bioquímicos que constituyen los seres vivos. Este proceso también se puede, por tanto, denominar como de asimilación del carbono.
La primera enzima que interviene en el ciclo y que fija el CO2 atmosférico uniéndolo a una molécula orgánica (ribulosa-1,5-bifosfato) se denomina RuBisCO (por las siglas de Ribulosa bisfosfato carboxilasa-oxigenasa).
Para un total de 6 moléculas de CO2 fijado, la estequiometría final del ciclo de Calvin se puede resumir en la ecuación:
6CO2 + 12NADPH + 18 ATP → C6H12O6P + 12NADP+ + 18ADP + 17 Pi
que representaría la formación de una molécula de azúcar-fosfato de 6 átomos de carbono (hexosa) a partir de 6 moléculas de CO2
Fases del Ciclo de Calvin
El ciclo está dividido en tres fases:1ª Fase: Fijación del CO2
La RuBisCO cataliza la reacción entre la ribulosa bifosfato (una pentosa, es decir un monosacárido de 5C, RuBP) con el CO2, para crear 1 molécula de 6 carbonos, la cual al ser inestable termina por separarse en 2 moléculas que contienen 3 átomos de carbono cada una, el fosfoglicerato (PGA). La importancia de la RuBisCo queda indicada por el hecho de ser el enzima más abundante en la naturaleza.2ª Fase: Reducción
Primero ocurre un proceso de activación en el cual una molécula de ATP, proveniente de la fase fotoquímica, es usada para la fosforilización del PGA, transformándolo en difosfoglicerato. Esa transferencia de un enlace fosfato permite que una molécula de NADPH+H+ reduzca el PGA, mediante la acción de la enzima gliceraldehído-3-fosfato-deshidrogenasa, para formar gliceraldehído-3-fosfato (PGAL). Esta última molécula es una triosa-fosfato, un azúcar de tipo aldosa con 3C, que es una molécula estable y con mayor energía libre (capaz de realizar mayor cantidad de trabajo) que las anteriores. Parte de PGAL se transforma en su isómero dihidroxiacetona-fosfato (cetosa de 3C). Estas dos triosas-fosfato serán la base a partir de la cual se formen el resto de azúcares (como la fructosa y glucosa), oligosacáridos (como la sacarosa o azúcar de caña) y polisacáridos (como la celulosa o el almidón). También, a partir de estos azúcares, se formarán directa o indirectamente las cadenas de carbono que componen el resto de biomoléculas que constituyen los seres vivos (lípidos, proteínas, ácidos nucleicos, etc.).3ª Fase: Regeneración
El ciclo continua a lo largo de una serie de reacciones hasta formar ribulosa-fosfato, que mediante el consumo de otra molécula de ATP, regenera la ribulosa bisfosfato (RuBP) original, dejándola disponible para que el ciclo se repita nuevamente.Por tanto, por cada vuelta del ciclo se incorpora una molécula de carbono fijado (CO2) a otra molécula preexistente de 5 átomos de carbono (ribulosa bisfosfato), el resultado final es la regeneración de la molécula de 5 átomos de carbono y la incorporación de un nuevo carbono en forma orgánica C(H2O). Para comprenderlo hay que tener en cuenta que el producto fundamental del ciclo de Calvin es el gliceraldehído-3-fosfato (de 3 átomos de carbono), molécula que sirve como base para la síntesis del resto de carbohidratos. Tras 3 vueltas del ciclo, una nueva molécula de PGAL sale de éste y puede ser posteriormente utilizada para la formación de otras moléculas.
Durante años se pensó que el ciclo de Calvin era independiente de la luz y se denominó "fase oscura de la fotosíntesis". Hoy en día se conoce perfectamente que tanto la actividad de RuBisCO como de otras enzimas clave del ciclo es regulada por la luz, desactivándose en condiciones de oscuridad y reactivándose en condiciones de iluminación.
SEMEJANZAS Y DIFERENCIAS ENTRE RESPIRACIÓN Y FOTOSÍNTESIS
DIFERENCIAS:
1) La Fotosíntesis se realiza en los CLOROPLASTOS y la Respiración se realiza en las MITOCONDRIAS.
2) La Fotosíntesis se realiza únicamente en las células con clorofila y la Respiración se realiza en todas.
3) La Fase luminosa de la Fotosíntesis se realiza en presencia de la LUZ y la Respiración se realiza tanto en la luz como en la oscuridad.
4) La Fotosíntesis transforma la energía luminosa en energía química y la Respiración transforma la energía química en calor y en energía aprovechable.
5) ) La fotosíntesis la realizan sólo los organismos AUTÓTROFOS, mientras que la respiración celular es realizada por todas las células de todos los seres vivos. La Fotosíntesis la realizan los Vegetales de color verde, algunas Bacterias, Euglenofitas y no la realizan los Animales. La Respiración la realizan todos los seres vivos.
6) La Fotosíntesis en células eucariotas, se realiza en cloroplastos; en procariontes, en membranas y carboxisomas. La Respiración en células eucariotas se lleva a cabo en las Mitocondrias y en Procariotas en los mesosomas.
7)En la fotosíntesis interviene la coenzima NADP mientras que en la respiración es la NAD.
8) Los Productos iniciales de la Fotosíntesis son el H2O, CO2 y sales minerales. Los Productos iniciales en la Respiración es la Materia orgánica. Los Productos finales de la Fotosíntesis son Materia orgánica y O2(se desprende). Los Productos finales de la Respiración son H2O y CO2(se desprende). La fotosíntesis produce GLUCOSA y O2, en la respiración, se produce CO2 y H2O.
9) La Fotosíntesis es un Proceso ENDERGÓNICO(y endotérmico). La Respiración es un Proceso EXERGÓNICO (y exotérmico) La Fotosíntesis es ANABÓLICA y la Respiración es CATABÓLICA.
SEMEJANZAS:
1) Los dos Procesos se producen en organulos celulares en las células eucariotas.
2) Necesitan enzimas y coenzimas para realizar ciertas reacciones.Son vias metabólicas.
3) Existen componentes inorgánicos como H2O, CO2, O2.
4) Hay materia orgánica.
5) Sintetizan ATP.
6) En la Fotosíntesis se produce Síntesis de ATP y en la Respiración también a nivel de sustrato y fosforilación oxidativa.
1) La Fotosíntesis se realiza en los CLOROPLASTOS y la Respiración se realiza en las MITOCONDRIAS.
2) La Fotosíntesis se realiza únicamente en las células con clorofila y la Respiración se realiza en todas.
3) La Fase luminosa de la Fotosíntesis se realiza en presencia de la LUZ y la Respiración se realiza tanto en la luz como en la oscuridad.
4) La Fotosíntesis transforma la energía luminosa en energía química y la Respiración transforma la energía química en calor y en energía aprovechable.
5) ) La fotosíntesis la realizan sólo los organismos AUTÓTROFOS, mientras que la respiración celular es realizada por todas las células de todos los seres vivos. La Fotosíntesis la realizan los Vegetales de color verde, algunas Bacterias, Euglenofitas y no la realizan los Animales. La Respiración la realizan todos los seres vivos.
6) La Fotosíntesis en células eucariotas, se realiza en cloroplastos; en procariontes, en membranas y carboxisomas. La Respiración en células eucariotas se lleva a cabo en las Mitocondrias y en Procariotas en los mesosomas.
7)En la fotosíntesis interviene la coenzima NADP mientras que en la respiración es la NAD.
8) Los Productos iniciales de la Fotosíntesis son el H2O, CO2 y sales minerales. Los Productos iniciales en la Respiración es la Materia orgánica. Los Productos finales de la Fotosíntesis son Materia orgánica y O2(se desprende). Los Productos finales de la Respiración son H2O y CO2(se desprende). La fotosíntesis produce GLUCOSA y O2, en la respiración, se produce CO2 y H2O.
9) La Fotosíntesis es un Proceso ENDERGÓNICO(y endotérmico). La Respiración es un Proceso EXERGÓNICO (y exotérmico) La Fotosíntesis es ANABÓLICA y la Respiración es CATABÓLICA.
SEMEJANZAS:
1) Los dos Procesos se producen en organulos celulares en las células eucariotas.
2) Necesitan enzimas y coenzimas para realizar ciertas reacciones.Son vias metabólicas.
3) Existen componentes inorgánicos como H2O, CO2, O2.
4) Hay materia orgánica.
5) Sintetizan ATP.
6) En la Fotosíntesis se produce Síntesis de ATP y en la Respiración también a nivel de sustrato y fosforilación oxidativa.
7) En ambos procesos intervienen las llamadas particulas F(CF
en cloroplastos),ATP sintetasas localizadas en las membranas internas,(tilacoides
en fotosíntesis,crestas en mitocondrias)
8) Ambos procesos necesitan de una Cadena transporte de electrones: En la Fotosíntesis (en fase luminosa) y en la Respiración (cadena respiratoria).
9) Formación pool C3 (triosafosfato) en la fase oscura y en la Respiración se produce en la glucólisis.
10) Se realizan en Eucariotas (La Fotosíntesis no se lleva a cabo en animales, hongos y algunos protozoarios) y Procariotas (La fotosíntesis solo en algunas bacterias).
TEMAS MICROBIOLOGÍA
-Características generales y estructura de los virus.8) Ambos procesos necesitan de una Cadena transporte de electrones: En la Fotosíntesis (en fase luminosa) y en la Respiración (cadena respiratoria).
9) Formación pool C3 (triosafosfato) en la fase oscura y en la Respiración se produce en la glucólisis.
10) Se realizan en Eucariotas (La Fotosíntesis no se lleva a cabo en animales, hongos y algunos protozoarios) y Procariotas (La fotosíntesis solo en algunas bacterias).
TEMAS MICROBIOLOGÍA
Los virus son formas acelulares que se replican independientemente de los cromosomas de
las células huésped, pero no independientemente de ésta. Necesitan de la maquinaria celular
para producir sus réplicas.
Los virus están constituidos por un ácido nucleico, una cápsida proteica,alguna otra proteína(o enzima) y en ocasiones presentan un envoltura membranosa.
La morfología vírica se basa en sus componentes y es la siguiente:
- Ácido nucleico: Pueden presentar ADN o ARN, pero nunca los dos simultáneamente. El
ácido nucleico puede ser a su vez monocatenario o bicatenario, lineal o circular.
- Cápisde: Protege al ácido nucleico y está constituida por la unión de proteínas globulares
denominadas capsómeros. La unión de éstos da lugar a la formación de tres tipos principales
de cápsidas:
- Icosaédrica: Es una forma poliédrica de 12 vértices, 20 caras triangulares y 30
aristas.
- Heliocoidal: Los capsómeros adoptan una disposición helicoidal, formando una
estructura tubular en cuyo interior se aloja el ácido nucleico.
- Compleja: La presentan los bacteriófagos, es decir, los virus que infectan a las
bacterias. La cápsida está constituida por dos partes, la cabeza y la cola. La
primera es helicoidal y aloja al ácido nucleico, mientras que la cola es una
estructura de fijación a la bacteria y de inyección del ácido nucleico en el
interior de ésta.
- Envoltura: La mayoría de las veces es de composición similar a la membrana plasmática de
las células eucariotas, generalmente procedente de la célula huésped a la que parasita.
Las características esenciales comunes a los ciclos de multiplicación de todos los virus
comprenden la entrada en el citoplasma de una célula hospedadora, la replicación de su
genoma para producir una descendencia de viriones, la liberación de éstos al medio
extracelular o ambiente y su superviviencia en él. En su reproducción todos los componentes
se sintetizan por separado y luego se ensamblan. Sin embargo, existen dos tipos de ciclo
vitales infectivos víricos: el ciclo lítico y el ciclo lisógenico.
Según mi opinión(y de otros autores) los virus son nanoorganismos pues cumplen las tres famosas funciones de un ser vivo:
-Nutrición: si entendemos nutrición como construcción a partir de material exógeno es evidente esa construcción en la nucleocápsida.
Relación: si entendemos por relación reacción al medio, es innegable la evolución de los virus(mutación) y por tanto esa relación.
-Reproducción: ésta función no la niega ningún autor pues es perfectamente entendible.

a)Las c.euc. no tienen dicha estructura(las c.e.v. tiene pared celular pero no bacteriana),no pudiendo acceder al interior ,si fuera el caso, para actuar sobre los ribosomas de mitocondrias y cloroplastos que son tipo 70S como los procarióticos,evidentemente no inhibirían los 80s del citoplasma.
b) Los virus no tienen pared ni ribosomas por lo que no les afectaría.Que se receten en una infección vírica se explica porque los virus debilitan el sistema inmune y las bacterias medran,para éstas serían los antib.
S19B4
Ciclo lítico de un virus
Los viriones (virus en fase extracelular) no realizan ninguna actividad fisiológica, por lo que no requieren sintetizar proteínas ni utilizan energía; son estructuras inertes. Así, el ácido nucleico viral se replica a expensas de la maquinaria y la energía de la célula infectada.
Existen dos sistemas de replicación de virus, el ciclo lítico y el ciclo lisogénico.
Así en los llamados virus bacteriófagos(infectan a bacterias):
Ciclo lítico
Se denomina así porque la célula infectada muere por rotura al liberarse las nuevas copias virales. Consta de las siguientes fases:
- Fase de adsorción o fijación: El virus se une a la célula hospedadora de forma estable. La unión es específica ya que el virus reconoce complejos moleculares de tipo proteico, lipoproteico o glucoproteico, presentes en las membranas celulares.
- Fase de penetración o inyección: el ácido nucleico viral entra en la célula mediante una perforación que el virus realiza en la pared bacteriana.
- Fase de eclipse: en esta fase no se observan copias del virus en la célula, pero se está produciendo la síntesis de ARN, necesario para generar las copias de proteínas de la cápsida. También se produce la continua formación de ácidos nucleicos virales y enzimas destructoras del ADN bacteriano.
- Fase de ensamblaje: en esta fase se produce la unión de los capsómeros para formar la cápsida y el empaquetamiento del ácido nucleico viral dentro de ella.
- Fase de lisis o ruptura: conlleva la muerte celular. Los viriones salen de la célula, mediante la rotura enzimática de la pared bacteriana. Estos nuevos virus se encuentran en situación de infectar una nueva célula.
J19A4
Ciclo del VIH,(de un retrovirus).

Cambia proteína S en vez de gp120 y células del ap. respiratorio en vez de linfocitos CD4 y tienes
más o menos el ciclo del covid-19.
J19B4

a) 0,5 puntos: fabricación de queso (o de yogurt…). Fermentación láctica (0,25 puntos cada una).
b) 0,5 puntos: anaerobiosis y en el citoplasma (0,25 puntos cada una).
c) 0,5 puntos: porque la glucosa se transforma en a. pirúvico primero y después en etanol y CO2 en la fermentación.
d) 0,5 puntos: por la producción de CO2 en la fermentación,éste se combina con la hemoglobina de la sangre produciendo axfisia.
S18A3
a) Defina que es un microorganismo. (0,5 puntos)
b) Cite dos ejemplos de microorganismos beneficiosos para el hombre. Razone la respuesta. (0,5
puntos)
c) Cite dos ejemplos de microorganismos patógenos en el hombre indicando el por qué. (0,5 puntos)
d) Defina el concepto de biotecnología microbiana. (0,5 puntos)
a)Microorganismos son seres vivos de tamaño microscópico que para ser observados es necesario
utilizar el microscopio óptico o electrónico. Pueden ser unicelulares o pluricelulares, procariotas o
eucariotas, autótrofos o heterótrofos (0,5 puntos).Según el contexto podrían incluirse los virus(actualmente considerados sólo entidades biológicas no seres vivos)
b) Beneficiosos para el hombre: levaduras y hongos (fermentaciones: vino, cerveza, pan, quesos….),
E.coli (microbiota intestinal),ésta puede tornarse perjudicial (0,5 puntos)
c) Perjudiciales: virus (gripe, sida, hepatitis, sarampión), bacterias (neumonía, sepsis, salmonella…),
hongos(pie de atleta,candidiasis), protozoo (malaria)…(0,5 puntos)
d) Biotecnología microbiana son procesos industriales que utilizan microorganismos o partes de ellos(como sus enzimas) como base para
obtener productos de utilidad para las personas, como medicamentos o alimentos. Los
microorganismos deben tener un crecimiento rápido, resistir el cultivo a gran escala y una producción
en gran cantidad y en el menor tiempo posible (0,5 puntos)
J18A5
a) ¿Qué tienen en común la fabricación de la cerveza y el pan? (0,5 puntos)
b) ¿Cuál es y de dónde procede la molécula de partida? ¿Cuál es y donde va la molécula resultante
de la reacción básica de estos procesos industriales? (1,25 puntos)
c) ¿Qué organismo es el responsable de esta reacción? (0,25 puntos)
a) Son productos de una fermentación alcohólica y utilizan el mismo microorganismo (0,5 puntos)
b) La molécula de partida es la glucosa que procede de la hidrólisis del almidón que contiene p.e el trigo en el pan y p.e de la cebada en el caso de la cerveza. Las moléculas resultantes son el etanol, que en el caso del pan se evapora por el calor del horno, y el CO2 que produce su esponjosidad.En el caso de la cerveza la maltosa procedente del almidón se transforma primero en glucosas y éstas en el etanol convirtiendo dicho producto en una bebida alcohólica y el CO2 (tufo)dará las "burbujas" tambíen se transformará en ácido carbónico dándole acidez.(corregido el sabor con diversas sustancias como el lúpulo)
c) Se trata de una levadura del género Saccharomyces cerevisae. (0,25 puntos)
J18B5
La penicilina bloquea la formación de la pared celular bacteriana. (2 puntos)
a) ¿Por qué esta alteración provoca la muerte de la bacteria? Razone la respuesta. (1 punto)
b) Cite los componentes estructurales de una bacteria (1 punto)
a) La alteración de la pared
bacteriana provoca la muerte celular ya que inhibe su función más importante, que es la de mantener
su forma celular frente a cambio osmóticos,otras funciones son la de otorgarle capacidad antigénica(es lo que estimula al s. inmune en una infección) y regular intercambio de sustancias(más la membrana plasmática). (1 punto)
b) Se clasifican en comunes y facultativos(f):Cápsula bacteriana(f), pared bacteriana, membrana plasmática, ribosomas, inclusiones,mesosomas,plásmidos,mucosas(f)(f) orgánulos especiales(f) ( vacuolas de gas, etc), cromosoma bacteriano, flagelos y pelos(f). (1 punto)
S17A5

a)Se trata de un esquema del ciclo lítico de un virus del tipo retrovirus.
b)

Fase 1 Acoplamiento y fusión con la membrana de la célula hospedadora(tb se puede decir adsorción y penetración),el material genético del virus,ARN(en este caso) está dentro(ha habido una descapsidación,la cápsida es ese "ataud de interior amarillo...").
Fase 2 Transcripción inversa del ARN a ADN.
Fase 3 Integración de éste ADN(ADNv=ADN vírico) en el de la célula infectada.
Fase 4 Transcripción del ADNv(al mismo tiempo que la célula hospedadora transcribe el suyo propio) en ARNmv(ARN mensajero vírico)
Fase 5 Traducción del ARNmv transcrito (la c. tb puede transcribir el suyo...) en proteínas víricas(Pv)
Las fases 2 a 5 se pueden llamar de mutiplicación y en ella otra denominada de "eclipsis" puesto que el material genético del virus(fuese ADN o ARN) no se ve.
Fase 6 Ensamblaje delARNv(que lleva toda la información del virus,es decir información para ser traducida a las Pv) y las Pv que forman la cápsida("estuche protéico" en el dibujo es el "ataud" en cuyo interior están ARNv y ciertas Pv).Tambíen hay otras Pv que no son de la cápsida p.e la enzima retrotranscriptasa y las que participarán en el acoplamiento(aquí son los "chupetes rojos).
Fase 7 Lisis(ruptura) de la célula hospedadora y liberación de los virus(o viriones)
c)Números: 1: representa la fase de adsorción, 2: la de penetración, 3: eclipse (3a retrotranscripción
o transcripción inversa, 3b: transcripción a ARN mensajero, 3c: traducción a proteína (síntesis de
proteínas víricas), 4: ensamblaje, 5: liberación. (0,8 puntos)
S17B5
El SIDA es una enfermedad que afecta a los humanos. El causante es un virus que tiene su material genético en forma de ARN. Cuando este virus infecta a una célula, puede permanecer latente durante años. (2 puntos)
a. ¿Qué tipo de ciclo reproductivo cree que presenta este virus? Justifique su respuesta. (1 punto)
b. ¿Cómo se llama al proceso que debe seguir el ARN del virus para integrarse en el ADN de la célula infectada? Explique brevemente en qué consiste. (1 punto)
a)Se trata de un ciclo lisogénico(de un retrovirus), así en la fase de ECLIPSE el ácido nucleico viral se inserta en el ADN de la célula hospedadora(antes el ARNv se tuvo que retrotranscribir) y permanece sin expresarse(transcribirse por lo tanto tampoco traducirse) un tiempo indefinido(por lo tanto no hay lisis y no es un ciclo lítico).El VIH es un lentivirus,los sujetos infectados pueden estar en este estado durante años al final de los cuales revertir el c. lisogénico en lítico y el sujeto desarrollar los signos y síntomas del SIDA.
b)Retrotranscripción (o Transcripción Inversa). El ARN del virus debe pasar a ADN para poder integrarse al ADN de la célula hospedadora. Un enzima hace una copia del ARN en forma de ADN ,dicha enzima(retrotrascriptasa o transcriptasa inversa) está porque el ARNv lleva la información para su síntesis(traducción,una enzima es esencialmente proteína).
J17A4

a) A (virus animal) y B (bacteriófago). (0,5 puntos) el segundo infecta a las bacterias.
b) (0,75 puntos)
b.1. A1 (ácido nucleico), A2 (cápside) y A3 (envuelta). B1 (cabeza), B2 (ADN) y B3 (cola helicoidal). (0,5 puntos)
b.2. Cápside (o cápsida)es la cubierta proteica que envuelve el genoma vírico(cada uno de las unidades que la forman se llaman capsómeros. (0,25 puntos)
c) Virus parásitos intracelulares obligados porque no pueden vivir fuera del huésped ni pueden
cumplir las funciones vitales de un ser vivo(si lo consideramos como tal) por si solos ,así un virus extracelular (virión) es inerte cambiando esta condición al encontrar una célula que infectar (0,75 puntos)




No hay comentarios:
Publicar un comentario